Бамбергский барьер
- 1 year ago
- 0
- 0



Гема́тоэнцефали́ческий барье́р (гемато-энцефалический барьер, ГЭБ) (от др.-греч. αἷμα , род. п. αἵματος — «кровь» и др.-греч. ἐγκέφαλος — «головной мозг») — физиологический гистогематический барьер между кровеносной системой и центральной нервной системой . ГЭБ имеют все позвоночные .
Главная функция ГЭБ — поддержание гомеостаза мозга. Он защищает нервную ткань от циркулирующих в крови микроорганизмов , токсинов , клеточных и гуморальных факторов иммунной системы , которые воспринимают ткань мозга как чужеродную. ГЭБ выполняет функцию высокоселективного фильтра, через который из артериального русла в мозг поступают питательные, биоактивные вещества; в направлении венозного русла с глимфатическим потоком выводятся продукты жизнедеятельности нервной ткани.
Вместе с тем, наличие ГЭБ затрудняет лечение многих заболеваний центральной нервной системы , так как он не пропускает целый ряд лекарственных препаратов .


Первое свидетельство о существовании ГЭБ было получено в 1885 году Паулем Эрлихом . Он обнаружил, что введённый в кровеносное русло крысы краситель распространился по всем органам и тканям, кроме мозга . В 1904 году он высказал неверное предположение о том, что краситель не проникает в ткань мозга при внутривенном введении, так как не имеет к ней сродства . Южноафриканский хирург (1862—1913), ученик Эрлиха, обнаружил в 1909 году , что введённый внутривенно краситель трипановый синий не проникает в ткань мозга, но окрашивает сосудистое сплетение его желудочков . В 1913 году он показал, что краситель, введенный в спинномозговую жидкость собаки или лошади, проникает в ткань головного и спинного мозга, а периферические органы и ткани при этом не окрашиваются . На основании этих опытов Гольдман предположил наличие барьера между мозгом и кровью, который задерживает нейротоксические вещества .
В 1898 году венские патологи (1869—1933) и (1868—1932) показали, что при введении жёлчных кислот в кровеносное русло нейротоксический эффект не возникал, однако при инъекции непосредственно в ткань мозга развивалась кома . Немецкий невропатолог Макс Левандовский повторил опыты Бидля и Крауса с гексацианоферратом калия . Получив схожие результаты, он впервые использовал термин «Blut-Hirn-Schranke» ( перегородка между кровью и мозгом , 1900), принятый впоследствии также и в англоязычной литературе ( blood-brain barrier ) .
В 1915 году швейцарский нейроанатом Константин фон Монаков в Цюрихе предположил, что хориоидное сплетение и нейроглия выполняют барьерную функцию. В последующие годы им совместно с сотрудниками было опубликовано несколько сугубо гистологических трудов, посвящённых хориоидному сплетению, которое один из его учеников ( чилийский психоаналитик Фернандо Альенде-Наварро, 1890—1981) в публикации 1925 года именует «экто-мезодермальным барьером» ( фр. barrière ecto-mésodermique ).

Термин «гематоэнцефалический барьер» ( фр. barrière hémato-encéphalique ) был введён в научный обиход швейцарским , а затем советским физиологом Линой Соломоновной Штерн (первой женщиной — членом Академии наук СССР ) в совместном со своими студентами (1888—1972) и Раймондом Готье (1885—1957) сообщении Женевскому медицинскому обществу (Société de Biologie et Médecine) за 21 апреля 1921 года :
Между кровью, с одной стороны, и спинномозговой жидкостью, с другой, есть особый аппарат или механизм, способный просеивать вещества, обыкновенно присутствующие в крови или случайно проникшие в неё. Мы предлагаем называть этот гипотетический механизм, пропускающий одни вещества и замедляющий или останавливающий проникновение других веществ, гематоэнцефалическим барьером.
Первые сообщения Лины Штерн и Эрнеста Ротлина на заседании Société de physique et d’histoire naturelle de Genève и их публикация в Schweizer Archiv für Neurologie und Psychiatrie о наличии защитного барьера между мозгом и кровяным руслом относятся к 1918 году . Штерн и Ротлину посредством тончайшей канюли удалось ввести 1 мг кураре в пространство четвёртого желудочка экспериментального животного и зафиксировать медленную диффузию нейротоксина из спинномозговой жидкости сквозь лептоменингиальные мембраны в глубокие ядра мозжечка . В 1921 году выходит первая обзорная статья Л. С. Штерн в Schweizer Archiv für Neurologie und Psychiatrie, а в 1923 году её влиятельная работа «La barrière hémato-encéphalique dans les conditions normales et pathologiques», включённая в двухтомный коллективный сборник, посвящённый 70-летию Константина фон Монакова (1853—1930) и изданный тем же журналом. В последнем обзоре, помимо обобщения экспериментальных и гистологических исследований ГЭБ, его роли в нормальной физиологии и нейропатологии, Штерн также рассматривает и его роль в фармакодинамике и фармакокинетике нейротропных препаратов. В последующие годы Штерн, основываясь на анализе обширного экспериментального материала, сформулировала положения о ГЭБ и определила его значение для деятельности центральной нервной системы . В 1935 году под её редакцией был опубликован первый коллективный сборник, целиком посвящённый данной теме («Гемато-энцефалический барьер», М.—Л.: Биомедгиз, 1935). За исследования гематоэнцефалического барьера Л. С. Штерн в 1943 году была награждена Сталинской премией , денежную составляющую которой она передала на строительство санитарного самолёта.
В 1930-х годах было сформулировано различие между гематоэнцефалическим и гематоли́кворным барьером .
Морфологические структуры, ответственные за ГЭБ, были детально изучены в 1960-х годах методами электронной микроскопии .
Масса головного мозга человека составляет приблизительно 2 % от массы его тела. При этом потребление кислорода центральной нервной системой составляет 20 % от общего потребления кислорода организмом. Также, в противоположность другим органам, мозг обладает наименьшими запасами питательных веществ. Нервные клетки не могут обеспечить свои энергетические потребности путём одного лишь анаэробного гликолиза . Прекращение поступления крови к мозгу в течение нескольких секунд приводит к потере сознания, а через 10 минут наступает гибель нейронов . Такие энергетические потребности головного мозга обеспечиваются за счёт активного транспорта кислорода и питательных веществ через ГЭБ .
Нормальное функционирование мозга возможно также лишь в условиях электролитного и биохимического гомеостаза . Колебания pH , концентрации калия в крови и других показателей не должны влиять на состояние нервной ткани. Циркулирующие в кровеносном русле нейромедиаторы не должны проникать в нервную ткань, где они могли бы изменить активность нейронов . Также мозг должен быть защищён от попадания в него чужеродных агентов, таких как ксенобиотики и патогенные микроорганизмы . ГЭБ — это также и иммунологический барьер, так как он непроницаем для многих микроорганизмов, антител и лейкоцитов .
Система кровеносных сосудов центральной нервной системы имеет ряд структурно-функциональных особенностей, отличающих их от сосудов других органов и тканей. Эти особенности обеспечивают функции питания, выведения продуктов жизнедеятельности и поддержания гомеостаза .
Нарушения ГЭБ могут вызывать поражения центральной нервной системы. Целый ряд неврологических заболеваний напрямую или косвенно связан с повреждением ГЭБ .



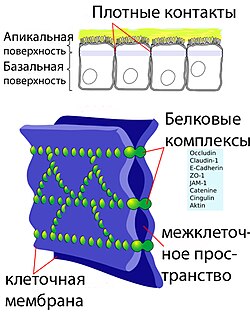
Основным элементом структуры ГЭБ являются эндотелиальные клетки . Особенностью церебральных сосудов является наличие плотных контактов между эндотелиальными клетками. В структуру ГЭБ также входят перици́ты и астроци́ты . Межклеточные промежутки между эндотелиальными клетками, перицитами и астроцитами нейроглии ГЭБ меньше, чем промежутки между клетками в других тканях организма. Эти три вида клеток являются структурной основой ГЭБ не только у человека, но и у большинства позвоночных .
Капиллярные сосуды выстланы эндотелиальными клетками. Эндотелий сосудов большинства тканей содержит открытые промежутки (фенестра́ции) диаметром около 50 нм и межклеточные щели от 100 до 1000 нм. Через эти промежутки вода и растворённые в ней вещества циркулируют между кровью и межклеточным пространством. Отличительной особенностью сосудов центральной нервной системы является отсутствие как фенестраций, так и межклеточных щелей между эндотелиальными клетками . Таким образом, эндотелиальная выстилка капилляров мозга является сплошной .
Другим отличием эндотелия церебральных капилляров от периферических является низкое содержание в них пиноцито́зных пузырьков (вези́кул) .
Количество митохондрий в эндотелиальных клетках сосудов мозга в 5-10 раз выше, чем в эндотелии периферических сосудов. Столь высокое содержание митохондрий связано со значительными энергетическими потребностями эндотелиальных клеток ГЭБ, осуществляющих активный транспорт и обмен веществ . (Митохондрии — это органеллы , в которых происходит синтез молекул АТФ , являющихся основным источником энергии для клеток.)
ГЭБ является также метаболическим или ферментативным (энзиматическим) барьером . На поверхности клеточных мембран эндотелиальных клеток ГЭБ находится целый ряд ферментов, причём в значительно большем количестве, чем на мембранах других клеток паренхимы . Это такие ферменты, как гамма-глутамилтрансфераза и фосфатаза (в частности глюкоза-6-фосфатаза), катехол-О-метилтрансфераза, моноаминоксидаза и цитохром Р450 . Благодаря высокой концентрации ферментов в эндотелиальных клетках ГЭБ многие вещества метаболизируются при транспортировании через цитоплазму этих клеток . Высота (размер в направлении, перпендикулярном стенке сосуда) эндотелиальной клетки ГЭБ составляет от 3 до 5 мкм (для сравнения, высота энтероцитов , эпителиальных клеток кишечника , - 17-30 мкм)

Соотношение холестерина к фосфолипидам в эндотелиальных клетках ГЭБ такое же, как и в эндотелиальных клетках периферических сосудов, и составляет ≈ 0,7 . Пассивный транспорт через клеточные мембраны ГЭБ происходит так же, как и пассивная диффузия в других эндотелиальных клетках . В мембранах эндотелиальных клеток содержится большое количество каналов, проницаемых для молекул воды. Они допускают диффузию воды между мозгом и кровеносной системой .
Благодаря отсутствию фенестраций и небольшому числу пиноцитарных везикул эндотелиальная выстилка капилляров мозга становится механическим барьером для крупных молекул и инородных веществ. Кроме этого, ГЭБ обладает значительным электрическим сопротивлением — около 1500—2000 Ом. (для сравнения, электрическое сопротивление для стенок капилляров мышечной ткани составляет лишь 30 Ом.)
Эндотелиальные клетки сосудов мозга плотно прилегают друг к другу. Между их стенками образуются так называемые плотные контакты, роль которых в обеспечении ГЭБ состоит в том, что они предотвращают проникновение в ткань мозга различных нежелательных веществ из кровеносного русла . Плотные контакты между эндотелиальными клетками блокируют межклеточный (парацеллюля́рный) пассивный транспорт . При этом блокируется парацеллюлярный транспорт веществ как из кровеносного русла в ткань мозга, так и в обратном направлении — из мозга в кровь .
Большое количество трансмембранных белков , таких как окклюди́н, разнообразные клауди́ны и замыкательные адгезионные молекулы связывают латеральные отделы клеточных стенок между собой, участвуют в формировании плотных контактов и делают возможным межклеточный транспорт и обмен веществ . Основными белками, обеспечивающими адгезию эндотелиальных клеток и формирование плотных контактов, являются клаудин-5 и клаудин-12 . Нокаут гена CLDN5, ответственного за синтез белка клаудина-5, приводил у экспериментальных мышей к тому, что их ГЭБ становился проницаемым для молекул с молярной массой до 800 г/моль. Такие генетически изменённые животные умирали через несколько часов после рождения .

Эндотелиальные клетки полностью покрывают подлежащий белковый слой, называемый база́льной мембраной . Толщина базальной мембраны колеблется от 40 до 50 нм. Она различима только под электронным микроскопом . Состоит в основном из коллагена IV типа, гепаринсульфат-протеоглика́нов, ламини́нов , фибронекти́на и других белков внеклеточного матрикса . Со стороны мозга базальная мембрана ограничена плазматической мембраной пластинчатых окончаний отростков астроцитов .

Перициты, ранее называвшиеся по имени первооткрывателя Шарля Мари Бенджами́на Руже́ (1824—1904) клетками Руже , являются составной частью ГЭБ . Они обладают несколькими важными для его функционирования свойствами: способностью к сокращению, регулированию функций эндотелия и макрофага́льной активностью .

Около 20 % поверхности эндотелиальных клеток церебральных капилляров покрыто относительно маленькими, овальными перицитами. Каждая 2—4-я эндотелиальная клетка имеет контакт с клеткой-перицитом . В основном перициты располагаются в местах контакта эндотелиальных клеток . Перициты имеются практически во всех артериолах, венулах и капиллярах организма. Уровень покрытия ими эндотелиального слоя капилляра коррелирует с проницаемостью сосудистой стенки. В органах и тканях с проницаемой сосудистой стенкой они могут мигрировать из кровеносного русла в межклеточное пространство. Так, например, в капиллярах скелетной мускулатуры соотношение перициты: эндотелиоци́ты составляет 1:100 .
Перициты, как и эндотелиоциты, располагаются на базальной мембране .
Также перициты синтезируют целый ряд вазоактивных веществ и играют важную роль в а́нгиогене́зе .
Перициты крепко связаны с эндотелиоцитами. Эта связь осуществляется благодаря трём типам контактов: щелевым соединениям , фокальным адгезиям и инвагинациям мембраны одной клетки в полость другой . Щелевые соединения непосредственно связывают цитоплазму двух клеток, являясь проницаемыми для ионов и небольших молекул . С помощью фокальных адгезий осуществляется прочная механическая связь двух типов клеток . Инвагинации участков цитоплазмы одной клетки в другую обеспечивают как механическое связывание, так и межклеточный обмен веществ .
Благодаря тесным контактам клетки опосредованно влияют на митотическую активность , экспрессию генов и, соответственно, фенотип друг друга .
Перициты содержат большое количество способного к сокращению белка актина . Благодаря этой своей структурной особенности они в состоянии изменять просвет капилляров и таким образом регулировать местное кровяное давление .
Данное свойство характерно только для церебральных перицитов. В капиллярной сети мозга они выполняют функцию макрофагов. Соответственно в цитоплазме церебральных перицитов располагается большое количество лизосом . В культуре тканей доказана способность перицитов к фагоцитозу и презентации антигенов .

Макрофагальные свойства перицитов образуют «вторую линию защиты мозга» от нейротоксических молекул , которые преодолели барьер эндотелиальных клеток . Таким образом они являются важной составной частью иммунной системы мозга. Сбой макрофагальной активности перицитов может стать одним из факторов развития целого ряда аутоиммунных заболеваний . Имеются данные об опосредованной роли перицитов в развитии болезни Альцгеймера .

Астроциты — большие нейроглиальные клетки звёздчатой формы. Своими отростками они выстилают стенки мозговых капилляров со стороны мозговой ткани. В то же время, несмотря на то, что пластинчатыми окончаниями их клеточных отростков выстлано около 99 % капиллярных сосудов, астроциты не выполняют прямой барьерной функции . Астроциты тесно взаимодействуют с эндотелиальными клетками. Между ними осуществляется постоянный обмен веществ . Астроглиальные клетки индуцируют возникновение и формирование ГЭБ. При проведении экспериментов по пересадке сосудов мозга в периферические органы и наоборот — периферических сосудов в ткань головного мозга, отмечено формирование ГЭБ в периферических сосудах, пересаженных в мозг (образование плотных контактов, перестройка эндотелиальных клеток), и разобщение эндотелиальных клеток и появление фенестраций между ними при пересадке мозговых сосудов . Также in vitro показано влияние астроцитов на фенотип эндотелия. В клеточной культуре, содержащей астроциты и эндотелиоциты, отмечено более плотное расположение эндотелия по сравнению с его чистой клеточной культурой .
Астроциты выделяют целый ряд веществ, которые влияют на проницаемость эндотелия . Эндотелиоциты в свою очередь выделяют ингибирующий лейкемию фактор (LIF), цитоки́н интерлейки́н-6 , которые воздействуют на процесс дифференциации астроцитов . Расстояние от пластинчатых окончаний отростков астроцитов до клеток эндотелия и перицитов составляет всего лишь 20 нм .
Главными задачами астроглиальных клеток является обеспечение нейронов питательными веществами и поддержание необходимой концентрации электролитов внеклеточного пространства . Астроциты синтезируют большую часть необходимого клеткам мозга холестерина . Холестерин не проникает через ГЭБ. В то же время в ткани мозга находится 25 % от общего холестерина организма. Бо́льшая его часть входит в состав миелина , который окутывает отростки нейронов аксоны . Нарушения процессов миелинизации нервных волокон вызывают развитие демиелинизирующих заболеваний, в частности рассеянный склероз .
Пластинчатые окончания отростков астроцитов неплотно покрывают со стороны мозга базальную мембрану сосудистой стенки с расположенными на ней эндотелиоцитами и перицитами. За счёт этого между эндотелиоцитами и тканью мозга возможна прямая диффузия различных веществ .
Заболевания, при которых происходит прямое или опосредованное поражение астроцитов (например, болезнь Альцгеймера , астроцитомы ), сопровождаются нарушением функционирования ГЭБ.
ГЭБ имеется в капиллярах большинства областей мозга, но не во всех. В циркумвентрикулярных органах ГЭБ отсутствует:
Данная гистологическая особенность имеет своё обоснование. Так например, нейрогипофиз выделяет в кровь гормоны , которые не могут пройти через ГЭБ, а нейроны дна IV желудочка ( лат. area postrema ) улавливают в крови наличие токсических веществ и стимулируют рвотный центр . Защитным барьером соседней с данными образованиями мозговой ткани является скопление таницитов . Они представляют собой клетки эпендимы с плотными контактами .
В среднем просвет капилляра мозгового сосуда составляет около 40 мкм . Наибольшая их плотность отмечена в коре головного мозга — от 300 до 800 капилляров на 1 мм³ ткани .
Суммарная поверхность стенок сосудов мозга составляет 12 — 20 м². Ежеминутно через сосудистую сеть мозга протекает около 610 мл крови со средней скоростью 1 мм/с создавая давление на её стенки 15-35 мм рт. ст. Через капиллярное русло мозга она проходит значительно быстрее (в среднем за 5 секунд), чем в других органах и тканях (для сравнения, в кишечнике , площадь сосудов которого достигает 180 м² среднее время прохождения крови ( англ. mean transit time ) равно 40 часам , а в печени с 70 м² — 30 секундам .
До конца XX века считалось, что у эмбриона и новорождённых ГЭБ не сформирован в полной степени и соответственно не выполняет своей функции. Причиной этого до сих пор широко распространённого мнения являются недостатки ранее проводившихся физиологических опытов. Эксперименты заключались во введении либо связанных с белками красителей, либо других маркеров взрослым животным и эмбрионам. Первые подобные опыты проводились в 1920 году . Маркеры, вводимые эмбрионам, проникали в ткань мозга и спинномозговую жидкость , в то время как у взрослых животных — нет. В ходе данных экспериментов был допущен ряд методических ошибок (использование чрезмерного объёма вводимого вещества, повышение осмотического давления ), из-за которых происходило частичное повреждение сосудистой стенки и соответственно маркер попадал в ткань мозга . При правильной постановке экспериментов пассажа маркера через сосудистую сеть отмечено не было .
В крови плода в большом количестве содержатся молекулы таких веществ как альбумин , α1-фетопротеин и трансферрин , отсутствуя при этом в межклеточном пространстве ткани мозга . В эмбриональном эндотелии обнаружен транспортёр Р-гликопротеин . Это свидетельствует о наличии ГЭБ в пренатальном периоде . В ходе развития организма происходит дальнейшее совершенствование ГЭБ .
Для небольших поляризованных молекул, например инулина и сахарозы , проницаемость ГЭБ эмбриона и новорожденного значительно выше, чем у взрослых . Схожий эффект отмечен и для ионов . Транспорт аминокислот и инсулина через ГЭБ значительно ускорен, по всей видимости, в связи с большой потребностью в них растущего мозга .
С другой стороны, в мозге эмбриона имеется дополнительный, отсутствующий у взрослых, барьер на границе между ликвором и тканью мозга — так называемые ремневы́е контакты ( англ. Strap Junctions ) между клетками эпендимы .
В ходе эволюции нервной ткани позвоночных происходит увеличение её объёма. Бо́льшая масса мозга требует лучшего обеспечения питательными веществами и выведения ненужных и отработанных веществ. Это привело к развитию густой капиллярной сети в ткани мозга. Следующим этапом эволюции стало появление защитного барьера от циркулирующих в крови токсичных для нейронов веществ — ксенобиотиков и токсинов .
У многих беспозвоночных ГЭБ отсутствует. У них эндотелий капилляров нервной ткани не образует сплошной выстилки сосудистой стенки. У высших беспозвоночных — насекомых , ракообразных и головоногих — защитный барьер между нейронами и кровью представлен исключительно глиальной тканью . В этом случае речь идёт о глиальном гематоэнцефалическом барьере .
У всех видов позвоночных имеется ГЭБ, и у большинства из них он образован преимущественно клетками эндотелия сосудистой стенки, скреплёнными между собой плотными контактами. Только у пластиножаберных (среди них акул и скатов ), а также семейства осетровых рыб ГЭБ формируется периваскулярными астроцитами. Из этого следует, что в процессе эволюции, вероятно, происходит расширение функций эндотелиальных клеток сосудов головного мозга, которые перенимают на себя барьерные функции.
Структурные различия глиального и эндотелиального гематоэнцефалических барьеров достаточно велики. Эндотелиальный барьер имеет целый ряд преимуществ. Одним из них является строгое разграничение функций эндотелиальных клеток и клеток астрогли́и, которые обеспечивают гомеостаз внеклеточной среды вещества мозга .
Кроме гематоэнцефалического барьера существует также гематоликворный, который отделяет центральную нервную систему от кровеносного русла. Он образован эпителиальными клетками с плотными контактами, выстилающими сосудистое сплетение желудочков мозга . Гематоликворный барьер также играет свою роль в поддержании гомеостаза мозга. Через него из крови в омывающую мозг спинномозговую жидкость поступают витамины , нуклеотиды и глюкоза . Общий вклад гематоликворного барьера в процессы обмена между мозгом и кровью невелик. Суммарная поверхность гематоликворного барьера сосудистых сплетений желудочков мозга приблизительно в 5000 раз меньше в сравнении с площадью гематоэнцефалического.
Кроме гематоэнцефалического и гематоликворного барьеров в организме человека существуют гематоплацента́рный , гематотестикуля́рный , , гематоретина́льный , и .


Гематоэнцефалический барьер не только задерживает и не пропускает целый ряд веществ из крови в вещество мозга, но и выполняет противоположную функцию — транспортирует необходимые для метаболизма ткани мозга вещества. Гидрофобные вещества и пептиды проникают в мозг либо с помощью специальных транспортных систем, либо через каналы клеточной мембраны. Для большинства других веществ возможна пассивная диффузия .
В капиллярах периферических органов и тканей транспорт веществ осуществляется в основном через фенестра́ции сосудистой стенки и межклеточные промежутки. В норме между клетками эндотелия сосудов мозга такие промежутки отсутствуют. В связи с этим питательные вещества проникает в мозг лишь через клеточную мембрану . Вода, глицерин и мочевина являются примерами тех небольших поляризованных молекул, которые могут свободно диффундировать через плотные контакты между эндотелиальными клетками ГЭБ .

Самой простой формой транспорта через ГЭБ является свободная (или пассивная) диффузия. Она может осуществляться как через клеточные мембраны эндотелиоцитов, так и через плотные межклеточные контакты. Для диффузии веществ движущей силой является разница концентраций. Диффузия веществ пропорциональна градиенту концентраций в кровеносном русле и ткани мозга. Для неё не требуется затрат клеточной энергии .
Липофи́льные структурные элементы клеточной мембраны, а также плотные межклеточные контакты снижают количество веществ, которые могут свободно диффундировать через ГЭБ. Проницаемость ГЭБ напрямую зависит от липофильности каждого конкретного вещества .
Проницаемость ГЭБ также зависит от молярной массы вещества. Молекулы с массой более 500 г/моль не могут диффундировать через ГЭБ. В то же время ГЭБ не является механическим барьером, который свободно пропускает молекулы меньшего размера и не пропускает большего. Процесс клеточной диффузии является динамическим, при этом он легче для веществ с молярной массой 200 г/моль, чем для веществ с 450 г/моль . Чем липофильнее и меньше вещество, тем легче оно диффундирует через клеточную мембрану .

Немецким биофизиком Германном Тро́йбле в 1971 году была высказана гипотеза о транспорте молекул с низкой массой через клеточную мембрану. Согласно ей они проникают в клетку через небольшие промежутки между цепями жирных кислот двойного слоя мембраны. Эти промежутки изменчивы, их образование не требует клеточной энергии . Теория Тройбле была спектроскопически доказана в 1974 году .
Прогноз и исследования проницаемости ГЭБ тем или иным веществом возможно проводить как in vitro так и in silico .
Липофильность и небольшая молекулярная масса не являются гарантией проницаемости ГЭБ для каждого конкретного вещества. Высокомолекулярные соединения (например, моноклона́льные антитела, рекомбина́нтные белки и другие) удерживаются ГЭБ .
Небольшие полярные вещества, например молекулы воды, с трудом могут диффундировать через гидрофобные отделы клеточной мембраны эндотелиоцита. Несмотря на это доказана высокая проницаемость ГЭБ для воды .
В клеточной мембране эндотелиоцита располагаются специальные гидрофильные каналы — аквапоры. В эндотелии периферических сосудов они образованы белком аквапорином-1 (AQP1), экспрессия которого ингибируется астроцитами в клетках сосудов мозга . На поверхности мембран клеток капиллярной сети мозга представлены в основном аквапорин-4 (AQP4) и аквапорин-9 (AQP9) .
Через аквапоры происходит регуляция содержания воды в веществе мозга. Они делают возможным быструю диффузию воды как в направлении мозга так и в направлении сосудистого русла в зависимости от осмотического градиента концентраций электролитов . Для глицерина , мочевины и ряда других веществ на поверхности клеточных мембран формируются собственные каналы — акваглицеропорины. В ГЭБ они представлены в основном белком аквапорином-9 (который также образует аквапоры) .

Процесс транспорта молекул через специализированные каналы осуществляется быстрее активного переноса с помощью специальных белков транспортёров. В то же время различные биологически активные вещества могут активировать или инактивировать транспортные каналы расположенные на клеточных мембранах .
Особой формой диффузии через клеточную мембрану является облегчённая диффузия. Целый ряд необходимых для мозга веществ, как например, глюкоза и многие аминокислоты, полярны и слишком велики для непосредственной диффузии через клеточную мембрану. Для них на поверхности клеточных мембран эндотелиоцитов располагаются специальные транспортные системы. Например, для глюкозы и аскорбиновой кислоты (витамина С) это GLUT-1-транспортёр. Их количество на поверхности обращённой в полость сосуда в 4 раза больше, чем на обращённой к мозгу.
Кроме транспортёров глюкозы на поверхности эндотелия располагаются множество белковых молекул выполняющих подобную функцию для других веществ. Так например MCT-1 и MCT-2 ответственны за перенос лактата , пирувата , мевалоновой кислоты , бутиратов и ацетатов . SLC7 транспортирует аргинин , лизин и орнитин . В геноме мыши выявлено 307 генов отвечающих за синтез SLC-белков, ответственных за облегчённую диффузию через клеточную мембрану различных веществ .
Транспортёры могут осуществлять перенос веществ в одном либо двух направлениях . В отличие от активного транспорта облегчённая диффузия направлена в сторону пространства (внутри- или внеклеточного) с меньшей концентрацией вещества и не требует затрат клеточной энергии.

В отличие от пассивного транспорта, не требующего затрат энергии, активный заключается в переносе веществ в пространство с большей концентрацией вещества и требует больших затрат клеточной энергии, получаемой при распаде молекул АТФ . При активном транспорте веществ из кровеносного русла в ткань мозга говорят о притоке вещества ( англ. influx ), в обратном направлении — об оттоке ( англ. efflux ).
В ГЭБ располагаются активные транспортёры энкефалина , антидиуретического гормона , [D-пеницилламин2,D-пеницилламин5]-энкефалина (DPDPE) .
Первым идентифицированным Efflux-транспортёром ГЭБ является Р-гликопротеин, который закодирован геном .
Впоследствии были открыты относящийся к классу ABC-транспортёров Multidrug Resistance-Related Proteine (MRP1) , Breast Cancer Resistance Proteine (BCRP) расположенный преимущественно на обращённой в просвет сосуда поверхности .
Некоторые Efflux- и Influx-транспортёры являются стереоселективными, то есть переносят лишь определённый стереоизомер (энантиомер) того или иного вещества. Так например, D-изомер аспарагиновой кислоты является прекурсором N-метил-D-аспартата (NMDA), который влияет на секрецию различных гормонов: лютеинизирующего гормона , тестостерона или окситоцина . L-изомеры аспарагиновой и глутаминовой кислоты являются стимулирующими аминокислотами и их избыток токсичен для ткани мозга . Efflux-транспортёр ASCT2 ( аланин - серин - цистеин -транспортёр) ГЭБ выводит в кровеносное русло L-изомер аспарагиновой кислоты, чьё накопление имеет токсический эффект. Необходимый для формирования NMDA D-изомер поступает в мозг с помощью других транспортных белков (EAAT, SLC1A3, SLC1A2, SLC1A6) .
В эпилептогенной ткани в эндотелии и астроцитах представлено большее количество белка Р-гликопротеина по сравнению с нормальной тканью мозга .
На клеточных мембранах эндотелиоцитов располагаются также транспортёры анионов (OAT и OATP) . Большое количество Efflux-транспортёров выводят из эндотелиоцитов целый ряд веществ в кровеносное русло .
Для многих молекул до сих пор не ясно выводятся ли они путём активного транспорта (с затратами клеточной энергии) или путём облегчённой диффузии .

С помощью рецептор-опосредованного трансцито́за происходит перенос больших молекул. На обращённой в просвет сосуда поверхности клетки расположены специальные рецепторы для опознавания и связывания определённых веществ . После контакта рецептора с веществом-мишенью происходит их связывание, участок мембраны инвагинируется в полость клетки и образуется внутриклеточный пузырёк — везикула . Затем она перемещается к обращённой к нервной ткани поверхности эндотелиальной клетки, сливается с ней и высвобождает связанные вещества. Таким образом во внеклеточное пространство мозга переносятся состоящий из 679 аминокислот белок трансферрин массой 75,2 кДа , липопротеины низкой плотности из которых образуется холестерин , инсулин и другие пептидные гормоны .
Одним из подвидов везикулярного транспорта является абсорбцио-опосредованный трансцитоз. Отмечается «прилипание» ряда положительно заряженных веществ ( катионов ) к отрицательно заряженной клеточной мембране с последующем образованием везикулярного пузырька и его переносом к противоположной поверхности клетки. Данный вид транспорта также называется катионным. Он проходит относительно быстрее рецептор-опосредованного трансцитоза .
Появление большого количества новых лекарственных веществ сделало изучение степени проницаемости ГЭБ для различных веществ крайне актуальным. Это относится не только к тем препаратам, которые используются в неврологии и нейрохирургии и чьё действие непосредственно зависит от их способности преодолевать ГЭБ, но и тем, которые используются в других областях медицины . Для исследования проницаемости ГЭБ применяется ряд методов. Классическим является проведение опытов на живых организмах ( in vivo ). Новые достижения науки сделали возможными эксперименты на клеточных культурах ( in vitro ), а также моделирование процесса на компьютере ( in silico ) . Результаты, полученные у млекопитающих ( in vivo ), могут быть использованы для описания проницаемости ГЭБ для того или иного вещества у человека.
Для определения проницаемости ГЭБ Ренкином (1959) и Кроне (1965) предложена модель, которая основывается на исследовании одного капилляра. Несмотря на свою упрощённость, она приближена к реальности . На основании данной модели определяется величина Кроне-Ренкина, которая показывает, какая часть вещества при прохождении через кровеносное русло мозга проникнет через ГЭБ . При её значении менее 0,2 ГЭБ слабопроницаем для вещества, при 0,2-0,8 — умеренно проницаем .
Симуляция процесса с использованием ЭВМ проводится в самых ранних фазах исследования. Высчитывается уровень свободной диффузии, учитывая ряд характеристик вещества: его липофильность, молярную массу, количество водородных связей и др.
Опыты in vitro проводятся для изучения транспортных процессов на клеточном уровне на изолированных капиллярах . В ходе эксперимента у подопытного животного выделяются сосуды. Обязательным является сохранение в них метаболической активности . Затем они помещаются между растворами с различными концентрациями исследуемых веществ. Молекулы могут быть маркированы. Метод позволяет определить проницаемость ГЭБ для конкретного вещества, а также процессы его переноса .
Первым, кто провёл in vivo исследования ГЭБ, был Пауль Эрлих. Эксперименты по проницаемости тех или иных веществ через ГЭБ заключаются в их непосредственном введении в кровеносное русло, а затем определении содержания в ткани мозга. По Вальтеру (F. Walter, 1929), вещества, применяемые с этой целью, должны удовлетворять следующим требованиям: распределяться в крови и цереброспинальной жидкости до того, как наступает их выделение, не расщепляться в организме и не связываться с белками; они не должны изменять состояние ГЭБ и приносить вред организму . Лишь при выполнении этих условий возможно определение проницаемости ГЭБ для определённого вещества in vivo .
Повреждения ГЭБ у человека наблюдаются при целом ряде заболеваний. Их коррекция рассматривается как терапевтическая стратегия .
Синдром дефицита белка GLUT-1 (G93.4 по международной классификации болезней ВОЗ ) — редкое аутосомно-доминантное наследственное заболевание, при котором отмечается нарушение синтеза белка GLUT-1, который ответственен за проницаемость ГЭБ для глюкозы и аскорбиновой кислоты . Заболевание проявляется в раннем детском возрасте. Недостаток поступления в ткань мозга глюкозы вызывает развитие микроцефалии , психомоторных нарушений, атаксии и целого ряда других неврологических расстройств .
Наследственная мальабсорбция фолиевой кислоты (D52.8 по международной классификации болезней ВОЗ ) — редкое аутосомно-рецессивное наследственное заболевание, при котором отмечается недостаток синтеза белка, обеспечивающего проницаемость ГЭБ для фолиевой кислоты.
Нарушение функционирования ГЭБ при болезни Альцгеймера приводит к увеличению количества амилоида β в мозге. Снижение количества спинномозговой жидкости приводит к повышению концентрации нейротоксичных веществ. Нейроваскулярная гипотеза патогенеза болезни Альцгеймера предполагает, что накопление амилоида β также связано с нарушением функционирования транспортеров, опосредующих перенос вещества из мозга в кровь, например, P-гликопротеина и LRP1 . При воспалительных процессах повышается захват амилоида β перицитами , что приводит к их гибели. Кроме того, при болезни Альцгеймера снижена эффективность транспорта инсулина через ГЭБ, играющего нейропротекторную роль .
Сахарный диабет (E10-E14 по международной классификации болезней ВОЗ ) является заболеванием, при котором возникает целый ряд функциональных и структурных изменений различных органов и тканей организма. Также отмечаются значительные изменения ГЭБ, которые проявляются в физикохимической перестройке мембраны эндотелиальных клеток и плотных контактов между ними .
См. также Хроническая цереброспинальная венозная недостаточность
Рассеянный склероз (G35 по международной классификации болезней ВОЗ ) — хроническое прогрессирующее заболевание нервной системы, при котором отмечается преимущественное поражение белка миелина ткани мозга.
Сосуды мозга здоровых людей непроницаемы для клеток крови, в том числе иммунных клеток. У больных рассеянным склерозом происходит миграция активированных Т-лимфоцитов в паренхиму мозга через ГЭБ, повышается уровень провоспалительных цитокинов — γ-интерферона, ФНО-α, ИЛ-1 и других; активируются В-лимфоциты. В результате начинают синтезироваться антитела к белку миелину, что приводит к формированию очагов воспалительной демиелинизации .

Ишемический инсульт (I63 по международной классификации болезней ВОЗ ) — острое нарушение мозгового кровообращения, обусловленное недостаточностью поступления крови к участкам центральной нервной системы.
Ишемический инсульт приводит к высвобождению оксидантов, протеолитических ферментов и цитокинов в ткани мозга, что в итоге вызывает развитие цитотоксического отёка и изменение проницаемости ГЭБ . В результате запускается процесс миграции лейкоцитов через эндотелий в ткань мозга, которые вызывают в том числе поражение здоровых клеток нервной ткани .
Лишь немногие попадающие в кровь патогенные микроорганизмы способны проникать через ГЭБ. К ним относятся менингококки ( лат. Neisseria meningitidis ), некоторые виды стрептококков — в том числе пневмококки ( лат. Streptococcus pneumoniae ), гемофильная палочка ( лат. Haemophilus influenzae ), листерии , кишечные палочки ( лат. Escherichia coli ) и ряд других. Все они могут вызывать воспалительные изменения как мозга — энцефалит , так и его оболочек — менингит . Точный механизм проникновения этих патогенов через ГЭБ до конца не изучен, однако показано, что воспалительные процессы оказывают влияние на этот механизм . Так, воспаление, вызванное листериями, может привести к тому, что ГЭБ становится проницаемым для данных бактерий. Прикрепившись к эндотелиоцитам капилляров мозга, листерии выделяют целый ряд липополисахаридов и токсинов , которые в свою очередь воздействуют на ГЭБ, делая его проницаемым для лейкоцитов. Проникшие в ткань мозга лейкоциты запускают воспалительный процесс, в результате которого ГЭБ пропускает и бактерии .
Пневмококки секретируют фермент группы гемолизинов, который образует поры в эндотелии, через которые и проникает бактериальный агент .
Менингококки и E. coli проникают через ГЭБ трансэндотелиально .
Кроме бактерий, через ГЭБ в ткань мозга могут проникать некоторые вирусы . К ним относятся цитомегаловирус , вирус иммунодефицита человека (ВИЧ) и Т-лимфотропный вирус человека (HTLV-1).
Внутримозговые опухоли головного мозга ( глиобластомы , метастазы в мозг и др.) выделяют целый ряд веществ , которые дезинтегрируют работу ГЭБ и нарушают его избирательную проницаемость. Такие повреждения гематоэнцефалического барьера вокруг опухоли может вызвать вазогенный отёк мозга .
ГЭБ избирательно проницаем для различных лекарственных веществ , что учитывается в медицине при назначении препаратов для лечения заболеваний центральной нервной системы (ЦНС). Такие препараты должны проникать в ткань мозга к клеткам-мишеням. Также имеет значение то, что при инфекционно-воспалительных заболеваниях ЦНС проницаемость ГЭБ повышается, и через него могут проходить те вещества, для которых он в нормальном состоянии служил непреодолимой преградой. Особенно актуально это для антибактериальных препаратов.
Проникновение антибактериальных препаратов через ГЭБ
| Хорошо | Хорошо при воспалении | Плохо даже при воспалении | Не проникают |
|---|---|---|---|
| Изониазид | Азтреонам | Гентамицин | Клиндамицин |
| Пефлоксацин | Амикацин | Линкомицин | |
| Рифампицин | Амоксициллин | Макролиды | |
| Хлорамфеникол | Ампициллин | Норфлоксацин | |
| Ко-тримоксазол | Ванкомицин | Стрептомицин | |
| Меропенем | Ломефлоксацин | ||
| Офлоксацин | |||
| Цефалоспорины III—IV поколения | |||
| Ципрофлоксацин | |||
| Левофлоксацин |