

Птицы Эдема
- 1 year ago
- 0
- 0

Пти́цы ( лат. Aves ) — группа теплокровных яйцекладущих позвоночных животных, традиционно рассматриваемая в ранге отдельного класса . Хорошо обособлена от остальных групп современных животных. Один из наиболее характерных признаков — покров из перьев , предохраняющий тело от неблагоприятных изменений температуры и играющий важную роль при полёте . Способность к полёту — важнейшая особенность птиц (отсутствие её у некоторых видов — вторичное явление) . Передние конечности приобрели форму крыльев . Птицы обладают особым строением органов дыхания и пищеварения , что тесно связано с их способностью летать. Ещё один отличительный признак птиц — наличие клюва .
Многие виды птиц совершают регулярные длительные миграции из одного региона Земли в другой. Ещё большее количество ведёт кочевой образ жизни, постоянно перемещаясь на небольшие расстояния в пределах своих ареалов. Птицы являются социальными животными, которые общаются между собой при помощи визуальных и звуковых сигналов и могут осуществлять общественные действия: коммунальное гнездование, совместную охоту или защиту от хищников . Для большинства видов характерна моногамия . Как правило, яйца откладываются в гнездо и насиживаются одним или обоими членами пары. Большинство птиц долго ухаживает за потомством после его появления на свет.
Птицы населяют все регионы земного шара, включая внутренние части Антарктиды . По данным Международного союза орнитологов , на октябрь 2023 года известно 11 001 вид ныне живущих птиц ( 19 818 подвидов), а также 160 видов, вымерших по вине человека после 1500 года . Это делает птиц наиболее разнообразным классом четвероногих . На территории России отмечено 789 видов, включая 657 гнездящихся видов , 125 видов, гнездование которых не показано, и 7 исчезнувших в стране видов .
Будучи наиболее многочисленной и широко распространённой группой высших позвоночных, а также вследствие особенностей своей биологии, птицы играют важную роль в природе и в жизни человека. Многие виды имеют важное экономическое значение. Птицеводство является одной из основных отраслей сельского хозяйства , производящей для человека мясо, яйца, жир и перья. Многие виды птиц содержатся человеком в качестве домашних животных . Вследствие деятельности человека многие птицы получили своё дальнейшее распространение, некоторые стали синантропами , но в то же время около 1200 видов в разной степени подвержены риску вымирания и охраняются национальными и международными законами .
Раздел зоологии , занимающийся изучением птиц, называется орнитологией .

Русское слово восходит к праслав. , от балтослав. «птица» + уменьшительного суффикса , от праи.-е. «маленький». Родственно аналогичным словам в славянских языках и латыш. «птица», лит. «маленькая птичка». Более дальнее родство — с лат. «цыпленок», санскр. «дитя, сын». Ложное родство — с греч. «птица» (от древнегреч. «летать»).
Латинское слово avis восходит к пра-и.е. *h 2 ewis (в традиционной реконструкции *awis ) «птица», от которого происходят также др.-инд. विः ( IAST : viḥ ) «птица», др.-греч. ἀετός «орёл», арм. հավ (hav) «курица», валл. hwyad «утка» .
Размеры и масса тела птиц очень различны. Самые мелкие птицы — колибри — обычно весят 3—7 г . Самые крупные летающие птицы обычно не превышают 12—16 кг по массе тела ( лебеди , грифы , дрофы ) и 3 м в размахе крыльев ( пеликаны , альбатросы , андский кондор ) , но иногда достигают массы 18—19 кг ( африканская большая и обыкновенная дрофы) и размаха крыльев 3,63 м ( странствующий альбатрос ) .
Крупнейшей известной летающей птицей всех времён был Pelagornis sandersi из олигоценовых отложений США: его размах крыльев оценивают в 6,1—7,4 м , а массу — в 20—40 кг . Примерно такой же размер имел аргентавис из миоцена Аргентины .
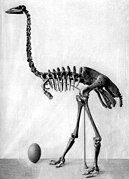
Нелетающие птицы могут быть намного тяжелее: крупные пингвины достигают 40 кг, казуары и страусы — 80—150 кг , а некоторые вымершие птицы ( эпиорнисовые , моа ) — 230—450 кг . В высоту большие моа достигали 3,7 м .
Самая маленькая современная птица — колибри-пчёлка : длина тела — около 5,7 см, а масса — около 1,6 грамма . Крупнейшая из ныне живущих птиц — африканский страус — ростом до 2,7 м и массой до 156 кг . Таким образом, самая маленькая птица современности примерно в 100 тысяч раз легче самой крупной.






Происхождение птиц долго оставалось предметом оживлённых дискуссий . За время исследований этой темы было выдвинуто несколько версий происхождения и родственных связей птиц и возникновения у них полёта, причём в течение более сотни лет они были чисто гипотетическими .
Впервые версия о происхождении птиц от рептилий возникла после открытия в 1861 году в Германии скелета археоптерикса — животного, жившего около 150 млн лет назад, в поздней юре . Он обладал признаками типичного пресмыкающегося — характерным строением таза и рёбер, зубами, когтистыми лапами и длинным, как у ящерицы , хвостом. Но на окаменелостях хорошо сохранились отпечатки маховых перьев , подобных таковым у современных птиц.
Долгое время археоптерикс оставался ископаемым животным, наиболее близким к современным птицам, поэтому историю класса представляли как эволюцию его потомков. На его изучении основывались все первые представления о происхождении птиц и их полёта: «древесная» («с деревьев вниз», Марш , 1880) и «бегательная» («с земли вверх», Уиллистон , 1879) теории . В соответствии с этим представляли и происхождение самих птиц — от триасовых текодонтов ( архозавроморф ) при древесной теории или от юрских бегающих тероподных динозавров при наземной .
К археоптериксу привязано и традиционное определение класса птиц, разделяемое многими авторами и ныне : «современные птицы, археоптерикс и все потомки их последнего общего предка» . Согласно ряду исследований, в традиционном классе птиц и даже в кладе авиал (птиц в широком смысле) археоптерикс — самый или один из самых древних и близких к общему предку известных представителей . Древнее и, возможно, базальнее него лишь несколько родов: Anchiornis , Xiaotingia , Aurornis и другие , но их не всегда относят даже к авиалам . С другой стороны, некоторые авторы приходили к выводу, что археоптерикс принадлежит не к авиалам, а к их сестринской группе , не дошедшей до наших дней, — дейнонихозаврам .
Впервые гипотезу о происхождении птиц от динозавров выдвинул в 1868 году Томас Гексли на основании сравнения строения археоптерикса и маленького хищного динозавра компсогната . С течением времени в связи с открытием многочисленных видов древних птиц и оперённых тероподовых динозавров, а также распространением кладистической классификации, большинство исследователей стало склоняться к тому, что птицы сами являются специализированными тероподовыми динозаврами из группы манирапторов , произошедшими от более ранних её представителей .
Согласно доминирующей в современной палеонтологии теории, птицы являются ныне живущими оперёнными динозаврами из клады теропод . Если быть точнее, птицы являются обособленной подгруппой манирапторов (клады теропод, наряду с птицами включающей в себя дромеозаврид и овирапторозавров , среди прочих). В связи с этим, всех остальных динозавров, кроме птиц, условно называют нептичьими динозаврами .
| Динозавры |
|
||||||||||||||||||||||||||||||||||||||||||||||||
Птиц роднит с динозаврами множество общих черт в строении скелета . По мере обнаружения всё новых видов теропод, граница между птичьими и нептичьими тероподами становится всё более размытой . Также если ранее одним из определяющих признаков птиц считалось наличие оперения , то ряд недавних находок в китайской провинции Ляонин показал, что перья имели и многие другие виды мелких теропод, например синозавроптерикс и синорнитозавр . Самым ранним из известных «перьев», вероятно, является позднеюрская окаменелость, описанная под названием Praeornis sharovi . Эволюцию перьев можно проследить начиная с таких крупных теропод, как карнотавр , покрытого многочисленными бугорками , похожими на фолликулы. Следующий этап виден у синозавроптерикса и Dilong paradoxus , покрытых волокнистым пухом . Наконец, у каудиптерикса , протархеоптерикса , синорнитозавра , микрораптора и Changyuraptor yangi уже были настоящие перья . В связи с этим было выдвинуто предположение о том, что все представители группы паравесов , а не только птицы, имели перья, жили на деревьях и были способны летать и/или планировать в воздухе . Тем не менее, предков птиц от других оперённых теропод отличал режим питания: тогда как предки птиц — авиалы , — были всеядны, остальные оперённые тероподы в основном питались мясом .
Ко времени возникновения полёта предки птиц были небольшими животными (относительно других динозавров). Судя по базальным представителям паравесов, их последний общий предок весил 600—700 г и имел длину около 65 см . Ранние птицы могли быть четырёхкрылыми, на что указывает четырёхкрылость некоторых ранних авиалов , троодонтидов и дромеозавридов (например, микрораптора ) . Это свидетельствует в пользу происхождения птиц от древесных планирующих форм, так как наземному бегающему животному длинные перья на ногах мешали бы .
Главным подтверждением теории эволюции птиц как клады динозавров является археоптерикс — теропод, представляющий собой переходную форму . Археоптерикс имел морфологические признаки и птиц (перья как у современных птиц), и рептилий (зубы, пальцы с когтями как у рептилий, а также длинный хвост как у ящерицы). Археоптерикс не является прямым предком птиц, но может быть тесно связан с истинным предком .
Наряду с теорией происхождения от динозавров существует гипотеза о происхождении птиц от более ранних архозавроморфов в поздней перми или раннем триасе . Раньше в качестве такого предка рассматривались мелкие и более примитивные псевдозухии из группы, традиционно называемой текодонтами (сейчас это таксономическое название устарело); теперь возможными предками считают преимущественно орнитозухид . Впервые эта теория была высказана в 1913—1914 годах Робертом Брумом и Фридрихом Хюне , позднее защищалась и оставалась наиболее популярной вплоть до конца двадцатого столетия. Тем не менее некоторые учёные, среди которых американские палеонтологи Ларри Мартин и , продолжают настаивать, что птицы произошли от более ранних архозавров типа лонгисквамы . Они утверждают, что сходство между птицами и манирапторами конвергентное . В конце 1990-х годов Федучия и Мартин выдвинули модифицированную версию своей гипотезы, согласно которой манирапторы являются нелетающими потомками ранних птиц . Эта гипотеза оспаривается большинством палеонтологов . В 2014 году в продолжение своей гипотезы Алан Федучия в соавторстве опубликовал исследование, в котором Scansoriopteryx рассматривается как древняя птица — потомок архозавров. По мнению авторов публикации, наличие у него архаичных черт и анатомических особенностей, характерных для птиц, и перьев на конечностях свидетельствуют о возникновении полёта «сверху вниз», при котором архозавры обзавелись перьями. Ими также утверждается, что Scansoriopteryx и все птицы произошли от архозавров, таких как склеромохлус . Филогенетические исследования никогда не подтверждали эту версию, и прежние аргументы авторов публикации ранее уже неоднократно были подвергнуты критике как ненаучные .
Ещё одну версию происхождения птиц отстаивал российский палеонтолог Евгений Курочкин . По его мнению, предком настоящих птиц мог быть протоавис , который на 75 млн лет древнее археоптерикса . Критики этой теории утверждают, что ископаемые остатки протоависа плохо сохранились и могут быть химерой .
В 1970-х годах палеонтолог после изучения скелета триасового текодонта- крокодиломорфа сфенозуха предложил гипотезу о происхождении птиц от общих с крокодилами предков в триасе. Она базировалась на большом количестве общих черт строения черепа ранних крокодиломорфов и птиц . Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной, поскольку сходство птиц и крокодиломорфов ограничивается типичными признаками архозавров.
Помимо описанных выше основных теорий происхождения птиц, были выдвинуты и многие другие, однако сегодня они не воспринимаются всерьёз. К таковым относятся версии о родстве птиц с ящерицами ( Фогт , 1879) или птерозаврами ( Оуэн , 1875; Сили , 1881), о происхождении птиц от эозухий ( Г. П. Дементьев , 1940) , о близости птиц и птицетазовых динозавров (Galton, 1970) или млекопитающих (Гардинер, 1982) .
| Кладограмма по Pei et al., 2020 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|

Более 40 % ключевых черт, обнаруженных у современных птиц, развилось в течение 60 миллионов лет перехода от самых ранних архозавров птичьей линии к первым манираптороморфам, то есть первым динозаврам. Утрата остеодерм , обычно характерная для архозавров, и приобретение примитивных перьев могли произойти на ранней стадии этой фазы. После появления манираптороморф следующие 40 миллионов лет ознаменовались непрерывным уменьшением размеров тела и накоплением неотенических (ювенильных) характеристик. Плотоядность становилась все менее распространённой, а размеры черепа и длина передних конечностей увеличивались. Покровы превратились в сложные перистые перья .
Самыми ранними известными представителями клады Avialae могут быть птицеподобные динозавры Anchiornis huxleyi , Xiaotingia zhengi , Aurornis xui и Eosinopteryx brevipenna , известные по окаменелостям из верхне юрской (около 160 млн лет назад) формации Тяоцзишань в Китае . Впрочем, их родственные связи малопонятны; согласно другим исследованиям, эти виды не входят в Avialae , а самые базальные известные представители группы — скансориоптеригиды , жившие примерно в то же время, археоптерикс , живший примерно на 10 млн лет позже, или другие таксоны . Эти ранние формы обладали необычными анатомическими особенностями, которые в ходе дальнейшей эволюции птиц были утеряны .
Эволюционное разделение птиц на множество разнообразных форм началось в меловом периоде . Многие из этих форм, в том числе и предки современных веерохвостых птиц , вначале сохраняли признаки ящеров, впоследствии редуцированные, — крылья с когтями и зубы. Джехолорнис и другие древнейшие птицы обладали длинным хвостовым отделом позвоночника . У представителей других, более развитых групп, например, клады пигостилевых , 4—6 задних хвостовых позвонков сливаются воедино, образовав пигостиль . Впрочем, это касается не всех пигостилевых (у большинства бескилевых , в том числе у некоторых летающих ( тинамуобразных ), хвостовые позвонки не срастаются) .

Первой широко распространённой и разнообразной группой короткохвостых птиц стали энанциорнисовые , или «противоположные птицы», названные так потому, что характер сочленения лопатки и коракоида у них противоположен таковому современных птиц . Энанциорнисовые не относятся к веерохвостым птицам и более примитивны, но были внешне похожи на них. В меловом периоде энанциорнисовые эволюционировали вместе с веерохвостыми и занимали различные экологические ниши : от обитателей песчаных морских побережий и охотников за рыбой до ведущих древесный образ жизни и питающихся семенами . В те же времена жили и птицы, ближе родственные веерохвостым: ихтиорнисы , которые напоминали современных чаек, тоже обитали на морских просторах и питались рыбой , а также гесперорнисообразные , которые настолько приспособились к охоте за рыбой, что утратили способность летать и полностью перешли к водному образу жизни .
Молекулярные часы показывают, что последний общий предок современных ( веерохвостых ) птиц жил ещё в мезозое . В пользу этого свидетельствуют и палеонтологические данные: уже в самом начале кайнозоя — в раннем палеоцене — существовали довольно далёкие друг от друга отряды пингвинообразных и совообразных . Некоторые исследователи относят к различным современным отрядам и ряд птиц позднемеловой эпохи . Но остатки этих птиц редки и обычно фрагментарны, состав позднемеловой орнитофауны остаётся плохо изученным, и степень диверсификации веерохвостых птиц в мезозое, как и время их появления, неясна .
Предки веерохвостых отделились от предков энанциорнисов не позднее раннего мела : в этой эпохе, около 130,7 млн лет назад, жила Archaeornithura meemannae , ближе родственная веерохвостым, чем энанциорнисам . Сами веерохвостые появляются в палеонтологической летописи в позднем мелу . Это вегавис , живший в самом конце мела, и, возможно, ряд более древних птиц, известных по фрагментарным остаткам. Ещё в мезозое веерохвостые разделились на две группы, известные сегодня как подклассы бескилевых и новонёбных . Бескилевые — небольшая группа (5 современных отрядов) нелетающих и плохо летающих птиц, а новонёбные включают всех остальных современных птиц (более 30 отрядов). В ходе эволюции новонёбных первым (среди дошедших до наших дней групп) обособился надотряд Galloanserae , объединяющий гусеобразных и курообразных . Время, когда произошло это разделение, дискуссионно — большинство специалистов считают, что основные группы веерохвостых обособились в меловом периоде, и ветвь Galloanserae отделилась от остальных новонёбных до мел-палеогенового вымирания . Другие исследователи полагают, что расхождение могло произойти уже после (и благодаря) гибели большей части животного мира . Разногласие отчасти вызвано различными методами исследований: молекулярные часы указывают на разделение основных групп в середине мелового периода, а анализ окаменелостей свидетельствует о начале палеогена . Это не удивительно, поскольку в ископаемом виде находят в основном организмы тех групп, которые уже успели значительно распространиться, причём для распознавания признаков определённой группы нужно, чтобы они были уже достаточно ярко выражены, а на момент разделения разных таксонов их представители практически неотличимы друг от друга. Исследование ископаемых птиц затруднено ещё и тем, что их тонкостенные кости обычно плохо сохраняются.

Примерно 60—40 млн лет назад произошла интенсивная адаптивная радиация веерохвостых птиц, возникло большинство современных отрядов. Окаменелости птиц этого времени малочисленны и не дают представления о темпах и характере эволюции. Птицы, известные из кайнозоя , вероятно, имеют общего предка выше уровня зубастых Hesperornithiformes и Ichthyornithiformes . Некоторые современные отряды новонёбных могли появиться ещё в мезозое: на это указывают остатки из кампанских и маастрихтских отложений Монголии и Северной Америки , которые могут принадлежать гусеобразным, пеликанообразным , курообразным, трубконосым , ржанкообразным и гагарообразным . Впрочем, эти остатки в основном фрагментарны и их идентификация дискуссионна .
Бескилевые , вероятно, произошли от общего предка, отделившегося от ранних килевых форм, сохранивших палеогнатическое нёбо . Форма килевой кости современных страусов [ уточнить ] предполагает, что их дальние предки летали, и что нелетающий образ жизни современных бескилевых птиц вторичен .
В большинстве отрядов птиц современные роды известны начиная со среднего или позднего миоцена . Многие из ныне живущих семейств можно проследить по ископаемым остаткам до позднего эоцена или олигоцена . Более ранние находки обычно настолько отличаются от современных родов, что установить их семейственную принадлежность часто затруднительно .
Наряду с современными отрядами в кайнозое возник ряд семейств и отрядов птиц, впоследствии вымерших . Среди бескилевых это гигантские нелетающие эпиорнисообразные и моаобразные , а также летающие литорнитообразные . С палеоцена по плиоцен в Южной Америке обитали фороракосовые — семейство очень крупных хищных нелетающих птиц из отряда кариамообразных . Ещё одну группу вымерших крупных нелетающих птиц составляют гасторнитиды из палеоцена и эоцена Северной Америки, Европы и Китая. Их выделяют в самостоятельный отряд Gastornithiformes , возможно, близкий к гусеобразным . С палеоцена по плиоцен на всех континентах жили гигантские летающие морские птицы из отряда Odontopterygiformes, примечательные ложными зубами и иногда превышавшие 6 м в размахе крыльев .

Первую попытку систематизации животных предпринял в IV веке до н. э. греческий учёный Аристотель — в своих сочинениях « О частях животных » и «О возникновении животных» он выделил всех известных ему птиц в «высший род» Ornithes . Несмотря на очевидное несовершенство этой системы, вплоть до второй половины XVII века новых попыток классификации животного мира не предпринималось. В 1676 году вышла в свет рукопись «Ornithologiae libri tres» , написанная английским биологом Френсисом Виллоуби , оформленная и опубликованная уже после его смерти . Это первая известная работа, в которой авторы попытались классифицировать птиц на основании внешних морфологических признаков. В 1713 году была опубликована одна из первых классификаций птиц, сделанная английским биологом Джоном Рэем . Дальнейшие основы классификации птиц были заложены в 1758 году Карлом Линнеем в « Системе природы », в которой он ввёл иерархические категории и биноминальную номенклатуру при обозначении видов, используемые вплоть до настоящего времени . В системе Линнея все животные были разбиты на шесть основных категорий — классов , — одну из которых заняли птицы, или Aves (позднее появились категории и более высокого порядка). В настоящее время название Aves обычно употребляется по отношению к группе, объединяющей всех потомков общего предка археоптерикса и современных птиц. Кладистическая систематика рассматривает группу Aves как кладу тероподных динозавров . Птицы и крокодилы — единственные современные представители архозавров .
К концу XIX века исследования Томаса Гексли , Михаила Мензбира , Макса Фюрбрингера , Ганса Гадова и других заложили основу для современных представлений о системе класса птиц. В отличие от своих предшественников, для анализа родства видов они использовали большой сравнительно-анатомический материал.
В дальнейшем многие авторы по сути совершенствовали имеющуюся систему птиц .
Современные птицы — чётко очерченная группа, резко отличающаяся от других позвоночных. Но среди ископаемых есть переходные формы , подобные археоптериксу, и поэтому возникает вопрос, где проводить границу класса. Традиционно в его состав включают современных птиц, археоптерикса и всех потомков их последнего общего предка. Современных птиц (точнее, всех потомков их собственного последнего общего предка) объединили в подкласс веерохвостых, или новых птиц (Neornithes) , а археоптерикса и ряд других ископаемых форм — в подкласс древних птиц ( Archaeornithes ), впоследствии вышедший из употребления. Веерохвостых, в свою очередь, делят на две группы — бескилевые и новонёбные . Их обычно рассматривают в ранге надотрядов , однако в работах Лайвези и Зуси — в ранге когорт .
Ряд последователей системы , таких как Жак Готье , и другие, распространяют название Aves лишь на веерохвостых птиц, что выводит за пределы класса Aves археоптерикса, энанциорниса , конфуциусорниса и ещё ряд ископаемых форм. Кладу , охватывающую все эти формы , — птиц в широком смысле — Готье назвал Avialae . Эти предложения были приняты не всеми авторами .
Современная филогения и классификация птиц ещё формируются . Сравнительный анализ анатомии птиц, ископаемых остатков и ДНК не привёл исследователей к единому мнению по этому вопросу.
К середине XX века существовал целый ряд сходных классификаций птиц, основанных преимущественно на данных сравнительной морфологии. Среди них наиболее распространённой была классификация Александра Ветмора , включавшая 27 современных отрядов. В ней автор разделял современных птиц на два крупных надотряда: Impennes (включавший пингвинов) и Neognathae (новонёбные, остальные современные птицы) .
Традиционной из современных морфологических систем также считается классификация по Клементсу ( Klements , 2007), претерпевшая три пересмотра. Согласно ей, веерохвостые птицы разделяются на 2 подкласса, объединяющие 33 отряда (6 из них только с ископаемыми формами) и 213 семейств (из них 42 — ископаемые) . Существует классификация по Howard & Moore (4 редакция — 2013), отличающаяся от системы Клементса выделением самостоятельных отрядов трёхпёрсткообразные (Turniciformes), удодообразные (Upupiformes) и Bucerotiformes .
В последние два десятилетия систематика и филогения птиц были значительно пересмотрены с широким внедрением различных методов молекулярного анализа . Первой классификацией птиц, основывающейся на использовании молекулярных биохимических данных, была классификация , построенная на сравнительном анализе данных ДНК-ДНК-гибридизации , которая была создана американцами Чарлзом Сибли и Джоном Алквистом в 1970-х годах. В 1990 году авторами была опубликована работа «Филогения и классификация птиц» ( Phylogeny and Classification of Birds ), посвящённая филогении и эволюции птиц на основе ДНК-ДНК-гибридизации. Однако классификация Сибли — Алквиста не стала общепринятой, морально устарев уже на момент своей публикации . В последующие два десятилетия появился ряд работ со своими вариантами молекулярной филогении птиц. На сегодняшний день имеется целый ряд схем филогении современных птиц, полученных по результатам различных молекулярных и генетических исследований. Многие выводы на основании их результатов согласуются с морфологическими и биогеографическими данными, а некоторые противоречат им. Филогении и классификации на основе результатов этих исследований, в зависимости от выбранных методик и маркеров, часто противоречат и друг другу (например, Hackett et al. , 2008; Pacheco et al. , 2011 ). Преимущественно это связано с несовершенством применяемых методик и трактовок их результатов, конкуренцией научных школ, ошибками в выборе маркеров и т. п. . Последний обширный молекулярный анализ филогении птиц Prum et al . (2015) по результатам секвенирования ДНК охватывает 198 видов современных птиц, представляющих все основные линии, а также 2 вида крокодилов в качестве аутгруппы. В нём применялись байесовский анализ и метод максимального правдоподобия , давшие хорошо обоснованные и идентичные филогенетические деревья для всех основных линий птиц. Результаты анализа времени дивергенции совпадают с палеонтологической летописью, поддерживая крупную радиацию птиц вскоре после мел-палеогенового вымирания .
Наряду с этими двумя направлениями развиваются компромиссные классификации, одновременно учитывающие последние достижения существующих молекулярных и морфологических классификаций птиц . Например, в 2001 году российским орнитологом Евгением Кобликом была предложена компромиссная система птиц, объединяющая традиционные морфологические представления и данные ДНК-ДНК-гибридизации. От классификации Ветмора она отличается широкой трактовкой страусообразных, выделением в отдельные отряды трёхперсток, фламинго , грифов Нового Света и рябков .
В России и большинстве других стран СНГ классификации на основании молекулярных данных не получили широкого распространения и большинство орнитологов пользуется вариантами классической системы, восходящей к Ветмору .
Международный союз орнитологов использует компромиссную классификацию по Gill & Wright (2006) . Она представляет собой версию классификации по Howard & Moore , модифицированную данными ДНК-исследований птиц, обобщёнными в работе « Avian Higher-Level Phylogenetics » (2003) . Согласно этой классификации, выделяют 40 отрядов, 252 семейства и 2359 родов птиц .
| Традиционная классификация по (2007) | Молекулярная классификация (1990) |
|---|---|
|
|

Большая общая подвижность, высокий уровень метаболизма и гомойотермия , развитие органов чувств, сложное поведение, способность приспосабливаться к меняющимся условиям среды, а также широкий спектр питания позволили птицам широко расселиться по всему земному шару. Птицы населяют практически все регионы Земли и все наземные биомы , включая внутренние районы Антарктиды. Например, снежный буревестник гнездится в глубине этого материка на расстоянии до 440 км от берега . По краям Антарктического щита местами гнездятся пингвины ( императорский , Адели ), гигантский буревестник , южнополярный поморник .
Населяют птицы и самые безводные пустыни и горы, вплоть до границы вечных снегов: на высотах до 4500 м гнездится бородач , однажды наблюдавшийся в полёте на высоте более 7300 м ; на Эвересте клушица была отмечена на высоте 7950 м . Во время миграций стаи пролетающих гусей и журавлей иногда наблюдали на высоте 7000—9000 м н. у. м. В 1973 году гриф Рюппеля столкнулся с самолётом над Кот-д’Ивуаром на высоте 11 277 м .
Заселяют птицы и наиболее удалённые от материков океанические острова. На вновь появившихся вулканических островах некоторые морские птицы поселяются ещё до появления там растений и других животных. Несколько семейств птиц приспособились к жизни на море, и некоторые виды морских птиц достигают берега только с целью размножения . Птицы обычно не ныряют глубже 50—60 м, но у королевского пингвина зафиксировано погружение на 318 м , а у императорского — на 564 м .
Каждому крупному географическому району свойственен характерный состав орнитофауны , включающий как эндемиков , так и виды, населяющие соседние области. Наибольшего биоразнообразия птицы достигают в тропических широтах: здесь встречается около 85 % ныне живущих видов и подвидов, тогда как в умеренных и холодных широтах — лишь 15 %. По мере удаления от тропиков к северным районам видовой состав орнитофауны беднеет . Многие семейства, характерные для тропических широт, либо совсем не проникают в умеренные широты, либо представлены там малым количеством видов . Больше всего эндемичных видов отмечено в следующих странах: Индонезия (397 эндемиков из 1531 вида местной орнитофауны), Австралия (355 из 751), Филиппины (183 из 556), Бразилия (177 из 1635), Новая Зеландия (150 из 287), Перу (109 из 1678). В России известно 13 эндемиков .
В пределах своего ареала каждый вид птиц распространён неравномерно (говорят о «кружеве ареала») . Существуют виды с небольшими ареалами либо же эндемики . Эндемизм у птиц характерен прежде всего для островных территорий и территорий, которые ограничены биотическими, климатическими или геологическими барьерами. Например, некоторые виды колибри свойственны лишь отдельным горам в Андах . Только на Ямале и Таймыре гнездится краснозобая казарка , а кулик- лопатень — только в приморской полосе Чукотского полуострова .
Вследствие интродукции человеком многие птицы обосновались в дикой природе регионов, где они никогда ранее не обитали. Во многих случаях такая интродукция была сознательной: например, обыкновенного скворца в конце XIX века завезли в Северную Америку . Иногда попадание в новую среду было случайным: так, содержащийся в неволе попугай-монах попал на волю и успешно обосновался в нескольких североамериканских городах . Ареал некоторых видов, таких как египетская цапля , химахима и розовый какаду , расширился далеко за пределы первоначального, так как сельскохозяйственное использование земель создало новую, подходящую для них среду обитания. С ростом городов увеличивается число мест с обильным и доступным кормом для синантропных видов птиц, которые неизбежно вступают в процессы синантропизации и урбанизации, приобретая ряд новых экологических особенностей и адаптаций .


Внешняя морфология птиц отражает их приспособленность к полёту . Существует относительно немного (около 60 видов и подвидов) нелетающих либо почти нелетающих птиц, в ходе эволюции утративших способность летать, которую имели их предки . Именно способность к полёту определяет особенности данного класса животных, включая общий план строения. Туловище птиц обтекаемой яйцеобразной формы, отличается компактностью .
В связи с тем, что полёт требует интенсивной мышечной работы, уровень обмена веществ у птиц чрезвычайно высок и потребность в пище велика: её дневная норма составляет 12—28 % общей массы тела . Температура тела у различных видов варьирует от 37 °C до 45,5 °C и в среднем у большинства видов составляет 42,2 °C . Более низкая температура тела свойственна веслоногим и поганкам (39,5°), страусу (37,8°), киви (37°) , а более высокая (до 45,5 °C ) характерна для мелких птиц . Температура тела у птиц значительно выше днём, нежели ночью (наоборот у сов), а у самок выше, чем у самцов . Такая высокая температура тела имеет большое физиологическое значение, являясь одним из важнейших условий высокой интенсивности обмена веществ и всех физиологических процессов в организме . Птицы не имеют потовых желёз . Охлаждение тела достигается при помощи высокоразвитой дыхательной системы. Птицы также охлаждаются, находясь в тени либо в воде. Некоторые виды с этой целью испражняются на свои ноги. Это известно у грифов, андского кондора и других стервятников, а также у некоторых голенастых .
Кожные покровы птиц тонкие, эластичные. В соединительнотканном слое имеются обильные пучки гладких мышц , прикрепляющиеся к очинам контурных перьев и изменяющие их положение. Единственная кожная железа птиц — копчиковая железа , которая находится над хвостовыми позвонками (отсутствует у бескилевых птиц , у некоторых дроф, голубей, попугаев и др.). Она производит маслянистый секрет, который птицы выдавливают клювом и смазывают им оперение, что способствует сохранению эластичности пера .
Все птицы имеют перьевой покров, тогда как у других современных животных он не встречается . Перья покрывают всё тело птицы, кроме клюва и дистальных частей задних конечностей. У некоторых птиц, как например у индеек и американских грифов , оперение на голове и шее либо отсутствует вовсе, либо выражено очень слабо . Перья растут на отдельных участках тела птиц, называемых птерилиями . Эти участки разделены другими, на которых перья не растут — аптериями. Лишь у немногих птиц, например, у пингвинов , перья растут равномерно по всему телу .

Строение пера: 1 — опахало, 2 — стержень, 3 — обычная поверхность, 4 — пуховая часть, 5 — очин.
|

Типы перьев у птиц: 1 — контурное, рулевое перо (перо хвоста), 2 — типичное маховое перо, 3 — покровное перо, 4 — нитевидное перо, 5 — кистеобразное перо, 6 — пуховое перо
|

Перья крыла птицы: 1 — маховые 1-го порядка; 2 — большие кроющие; 3 —
крылышко
; 4, 8 — маховые 2-го порядка; 5 — оперение плеча; 6 — средние кроющие; 7 — кроющие крыла
|
Начальные стадии эмбрионального развития пера сходны с развитием чешуи рептилий, но детали происхождения перьев неясны. Они могут быть производными чешуи или эволюционным новшеством на основе механизма её развития .
Контурные перья покрывают всё тело птицы и имеют хорошо развитый плотный стержень, основание которого — полый очин — охватывается находящейся в коже перьевой сумкой. От стержня отходят упругие бородки, несущие бородочки с крючочками, которые сцепляются с крючочками соседних бородок, образуя опахало пера. В самой нижней ( пуховой ) части опахала бородки обычно более мягкие и длинные, а их бородочки не имеют крючочков. Особенности строения перьев у разных групп птиц могут отличаться. Так, виды, живущие в суровых температурных условиях, обычно обладают более развитой пуховой частью опахала .
У большинства птиц имеются пуховые перья (стержень мягкий) и пух (стержень совсем редуцирован), мягкие и длинные бородки которых несут мягкие бородочки, лишённые крючочков, из-за чего сцепленного опахала не образуется. Пуховые перья обычно располагаются по птерилиям. Пух относительно равномерно покрывает всё тело (веслоногие, гусеобразные, многие хищные птицы и др.), либо имеется только на аптериях (цапли, куриные, совы, многие воробьиные и др.), либо — только на птерилиях ( тинаму ). Обычно пуховые перья и пух прикрыты контурными перьями. Лишь у немногих птиц ( грифы , марабу и др.) голова и часть шеи покрыты только пухом. Нитевидные перья расположены под контурными перьями, обладают длинным тонким стержнем и редуцированными бородками. Видимо, они выполняют осязательную функцию .
У козодоев и особенно у сов хорошо выражена бархатистость (ворсистость) всех перьев, в том числе маховых и рулевых; гася завихрения воздушных потоков, она обеспечивает бесшумность полёта. У водоплавающих птиц относительно короткие и сильно изогнутые перья плотно прилегают друг к другу, предотвращая намокание оперения; при этом его водостойкость обеспечивается и особой формой бородок и бородочек, способствующей возникновению на поверхности пера водной плёнки .
Общее число перьев у крупных видов больше, чем у мелких. Например, у колибри около 1000 перьев, у мелких воробьиных 1500—2500, у чаек до 6000, у уток 10 000—12 000, у лебедей 25 000. Общее число перьев, их размеры и особенности строения варьируют и у близких групп в зависимости от их экологической специализации .
Окраска перьев может быть обусловленной как пигментами, так и структурными особенностями. Наиболее распространены пигменты двух типов: меланины и липохромы . Меланины дают различные оттенки чёрного, коричневого, красновато-бурого и жёлтого цветов. Липохромы обеспечивают более яркую окраску: красную, зелёную, жёлтую, синюю и др. Белая окраска создаётся полным отражением света от наполненных воздухом прозрачных роговых клеток пера при отсутствии пигментов . Структурная (оптическая) окраска бывает радужной (изменяется в зависимости от угла зрения) и однотонной. Она обусловлена тонкослойной интерференцией света в роговых клетках перьев, что придаёт окраске бо́льшую яркость. Окраска перьев варьирует в зависимости от пола ( половой диморфизм ), возраста птицы, времени года и питания .
Функции оперения многообразны. Оно обеспечивает возможность полёта , образуя несущие плоскости (крылья, хвост), и создаёт обтекаемость тела. Перья защищают кожные покровы от механических повреждений. Очень эффективны водозащитная и теплозащитная функции оперения .

Единичное перо
|

Перо
павлина
|

Структура пера
сине-жёлтого ара
|

Попугай ара, чистящий оперение
|
Периодически птицы линяют. Линька обеспечивает замену изношенного оперения и изменение окраски в зависимости от времени года ( брачное оперение ). Обычно птицы линяют один раз в году, реже два, а совсем редко (например, морянки ) — три раза в год. Крупные хищные птицы способны линять раз в несколько лет. В случае повторной линьки различают гнездовой и зимний наряды, а также пуховой наряд для неоперившихся птенцов. Как правило, смена маховых и рулевых, необходимых для полёта, перьев происходит в определённой последовательности, и птицы в процессе линьки сохраняют лётные качества. Из этого правила есть исключения — например, у утиных все маховые опадают одновременно, в результате чего они временно теряют способность к полёту . Перед насиживанием у самок большинства видов на брюхе образуется наседное пятно — неоперённый участок кожи с развитыми кровеносными сосудами, которым птица прижимается к яйцам и согревает их .
Перья требуют тщательного ухода, и птицы ежедневно тратят в среднем около 9 % времени на чистку оперения, купание и пылевые ванны . Ухаживают за перьями птицы с помощью клюва. Лишь одна птица не может этого делать — у колибри-мечеклюва клюв настолько длинный, что ей приходится чистить перья не клювом, а лапкой . С помощью клюва из перьев удаляются чужеродные предметы, после чего тело покрывается жиром, выделяемым из копчиковой железы у основания хвоста. Выделения придают оперению эластичность и действуют как противомикробное средство . Птицы с редуцированной или отсутствующей копчиковой железой, например попугаи, дневные хищники, тукановые и др., пользуются при чистке оперения особой «пудрой» с пудре́ток — участков тела, покрытых особыми т. н. «порошко́выми перьями». Такие перья по мере роста постоянно обламываются и крошатся на микроскопические чешуйки, образуя мелкодисперсный порошок, которым птицы натирают перья, чтобы предотвратить их намокание и слипание . Наиболее развиты пудретки у семейства цаплевых .
Практически все виды птиц охотно купаются: стоя или приседая в мелкой воде, они распускают и трясут перьями, стараясь равномерно смочить их. Птицы с быстрым полётом — стрижи , ласточковые и крачки на лету могут окунаться в воду . Многие виды купаются в росе или воде, скопившейся на листьях и траве, а также в дождевой воде. Некоторые также принимают пылевые ванны . Ряд видов для избавления от паразитов или с другими целями копаются в муравейниках , нанося на тело муравьиную кислоту (« муравление ») .

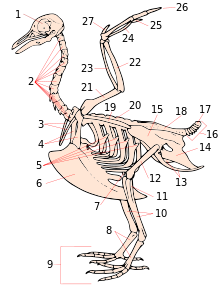
В строении скелета птиц отчётливо видны адаптации к полёту как основному способу передвижения, а у ряда видов — приспособления к нырянию, плаванию, бегу по земле или лазанию по стволам деревьев, скалам и другим вертикальным поверхностям .
Скелет птиц сильно упрощён и образован лёгкими и прочными костями. Некоторые кости имеют наполняемые воздухом полости, называемые « пневматическими », связанные с органами дыхания .
Кости черепа слиты воедино и не имеют черепных швов . Глазницы большие и разделены между собой костной перегородкой. Череп соединяется с позвоночником при помощи одного затылочного мыщелка , нижняя челюсть прикрепляется к черепу посредством квадратной кости , играющей роль подвеска. На голове вперёд выдаётся клюв, образованный надклювьем и подклювьем. Он покрыт роговым чехлом — рамфотекой . В проксимальной части надклювья у некоторых птиц (голуби, попугаевые , дневные хищные) есть восковица . Очень важная особенность черепа птиц — подвижность надклювья, которая увеличивает силу укуса и размеры зева, а подвижное нёбо помогает проталкиванию пищевого комка в пищевод. Форма и длина клюва, особенности рамфотеки, характер подвижности, размеры ротового отверстия у птиц варьируют в широких пределах, отражая пищевую специализацию видов .

Позвоночный столб делится на шейный, грудной, поясничный и хвостовой отделы. Для всех отделов позвоночника, кроме шейного, характерно сращение многих позвонков между собой. Это обусловливает жёсткость скелета туловища, что имеет большое значение при полёте. Шею образуют 9—25 шейных позвонков. Шейный отдел отличается большой гибкостью, однако подвижность позвоночника значительно уменьшается в грудном и полностью отсутствует в поясничном отделе . В грудном отделе 3—10 позвонков, которые у многих птиц срастаются, образуя спинную кость . Все поясничные, крестцовые и часть хвостовых позвонков вместе с тазом слиты в единую кость — сложный крестец . Число свободных хвостовых позвонков — от 5 до 9. Последние хвостовые позвонки у большинства видов срастаются в копчиковую кость ( пигостиль ).
Рёбра состоят из двух частей — позвоночной и грудинной. У птиц (кроме паламедей ) рёбра снабжены крючковидными отростками , к которым прикрепляются межрёберные мышцы . Грудина у летающих птиц и у пингвинов имеет киль , к которому прикрепляется мощная летательная мускулатура (подключичные и большие грудные мышцы); у бегающих птиц ( страусов , казуаров и др.) киля нет. Плечевой пояс состоит из мощных удлинённых коракоидов , сросшихся ключиц ( вилочки ) и длинных узких лопаток , которые лежат над рёбрами.
Передние конечности, превращённые в крылья , состоят из плеча (с плечевой костью), предплечья (с лучевой и локтевой костями) и кисти. Кисть, в отличие от плеча и предплечья, сильно преобразована по сравнению с рептильной: некоторые кости в ней редуцировались , а некоторые другие слились между собой. Кости пясти и некоторые кости запястья срастаются в пряжку (свободными остаются только две проксимальные косточки запястья). Сохранились рудименты трёх пальцев, среди которых наиболее развит средний. Передний палец несёт крылышко . В результате этих изменений передние конечности не используются при передвижении по земле .
Таз птиц открытый: лобковые и седалищные кости своими концами не соединяются внизу с соответствующими костями противоположной стороны (лобковое сочленение есть только у страусов), что позволяет откладывать крупные яйца .
Большая берцовая кость срастается с проксимальными костями предплюсны ( пяточной и таранной ) в одну кость — тибиотарзус . Малая берцовая кость частично редуцирована. Коленный сустав спереди прикрывает коленная чашечка, особенно хорошо развитая у ныряющих птиц . Дистальный ряд костей предплюсны и все кости плюсны срастаются в цевку , что даёт дополнительный сустав на ногах и увеличивает длину шага. Особенно удлинена цевка у околоводных птиц ( куликов , голенастых , журавлей ) .
Обычно ноги четырёхпалые (в преобладающем большинстве случаев три пальца направлены вперёд, а один — назад), но иногда число пальцев сокращается до трёх и даже двух (африканский страус) . У совообразных , бананоедов , скопы и некоторых других второй палец может произвольно быть направлен либо вперёд, либо назад. У ряда древесных птиц ( попугаи , кукушки , дятлы) два пальца направлены вперёд, два (первый и четвёртый) — назад . Когда птица садится, сгибание её ног под действием её веса приводит к натягиванию сухожилий , сгибающих пальцы ноги; при этом упомянутые сухожилия фиксируются в своих сумках благодаря поперечным рёбрышкам. Благодаря этому птицы держатся на ветках без мышечных усилий: пальцы крепко охватывают ветку, пока не будут задействованы мускулы-разгибатели .
Последняя фаланга каждого пальца ноги прикрыта хорошо развитым когтем. Слабо развитые рудиментарные коготки имеются у некоторых птиц на первом (некоторые хищные птицы, гуси , паламедеи ) и на втором пальцах крыла ( гоацин , казуары , киви ). У многих фазановых на цевке развиваются крупные костные выросты, покрытые роговым чехлом — шпоры .
Мускулатура птиц отличается большой плотностью, имеет тёмную окраску, а мышцы конечностей — длинные сухожилия. Очень развита мускулатура шеи в связи со сложностью движений головы. Наибольшего развития достигают грудные и подключичные мышцы, приводящие в движение крылья (у хорошо летающих птиц грудные мышцы составляют до четверти массы тела ). Сложно устроена мускулатура хвоста. Мышцы этой группы поднимают и опускают хвост и расправляют или сворачивают рулевые перья. Мощно развиты также мышцы ног, выполняющие большую работу при хождении птицы и передвижении по земле, веткам деревьев, во время взлёта и посадки .

Пищеварительная система птиц начинается ротовой полостью . Зубов у современных птиц нет — их отчасти заменяют острые края клюва , которым птицы захватывают, удерживают и порой размельчают пищу. Дно ротовой полости заполняется телом языка, весьма разнообразного по величине и форме в зависимости от характера питания. В ротовой полости располагаются слюнные железы . У некоторых птиц липкая слюна обеспечивает приклеивание мелкой добычи к языку (дятлы и др.). У видов, питающихся достаточно влажным кормом, слюнные железы слабо развиты или отсутствуют. У некоторых птиц под языком есть сильно растяжимый мешок, образованный стенками ротовой полости, играющий роль места временного хранения пищи ( пеликаны , кедровка , некоторые чистики ) .
Следующий за глоткой длинный пищевод у некоторых птиц ( куриные , дневные хищники, голуби, попугаи и др.) имеет чётко выраженное расширение — зоб , служащий для временного хранения пищи. В нём корм, подвергшийся обработке слюной, набухает и размягчается. Зобные железы некоторых птиц (например, голубей) вырабатывают особый творожистый секрет — « птичье молоко », который используется для выкармливания птенцов. У фламинго и пингвинов похожий секрет выделяют железы пищевода и желудка.
В связи с утратой зубов функция размельчения пищи перешла к желудку. Особенность птиц — разделение желудка на два отдела: железистый (где выделяются пищеварительные ферменты) и мускульный. Из пищевода пища попадает в железистый отдел, а оттуда — в мускульный. Стенки последнего образованы мощными мышцами, а в полости обычно находятся гастролиты . Они и складки желудка при сокращении мышц стенок перетирают и размельчают содержимое, компенсируя отсутствие зубов . Не поддающиеся перетиранию компоненты (шерсть, перья, хитин , осколки костей и т. п.) у многих птиц ( совы и дневные хищники, чайки, некоторые воробьиные и др.) спрессовываются в мускульном желудке в погадку и отрыгиваются наружу .
Кишечник относительно короткий: у разных видов он длиннее туловища в 3—12 раз, редко больше. Относительно длинный он у видов, питающихся грубой растительной пищей. В нём выделяют более длинный тонкий отдел и менее протяжённый толстый отдел. Слепые кишки обычно парные, у большинства птиц малы и выполняют роль лимфоидных органов. Однако у некоторых растительноядных видов (страусы, куриные, многие гусеобразные и др.) слепые кишки имеют большие размеры, и в них идёт активное переваривание пищи . Прямая кишка у птиц не развита, в связи с чем фекалии не накапливаются в кишечнике, облегчая массу птицы. Кишечник заканчивается расширением — клоакой, в которую также открываются мочеточники и протоки половых желёз .
Печень у птиц относительно велика, имеет вид двух лопастей и располагается в поперечном направлении в правой половине тела .
Для компенсации энергозатрат во время полёта требуется высокий обмен веществ . Поэтому у большинства птиц пищеварительная система адаптирована к быстрому перевариванию . Пищеварение у птиц протекает очень быстро: у свиристеля ягоды рябины проходят весь кишечник за 8—10 минут , а у утки через 30 минут после проглатывания карася длиной 6 см его остатков уже нельзя обнаружить в кишечнике . Энергетические резервы накапливаются в виде гликогена в печени и особенно в виде жира — подкожного и внутреннего. В период миграций запасы жира у некоторых видов могут достигать 30—50 % от общей массы тела .
Как и пресмыкающиеся, птицы обычно урикотеличны — их почки извлекают азотсодержащие отходы из кровотока и выделяют их в виде мочевой кислоты , а не мочевины или аммиака , как у млекопитающих. Крупные метанефрические почки лежат в углублениях тазового пояса. Мочеточники открываются в клоаку, мочевой пузырь отсутствует . Некоторые птицы, как например колибри, составляют исключение — их азотистые отходы могут выделяться в виде аммиака, то есть эти птицы по сути аммонотеличны . Ещё одним конечным продуктом обмена веществ является креатин (у млекопитающих эту функцию выполняет креатинин ) .
Моча из почек и экскременты из кишечника смешиваются и затем выделяются из клоаки птицы .
У морских птиц ( трубконосые , чайки , чистики , пеликанообразные и др.) и некоторых пустынных птиц ( африканский страус , пустынная курочка и др.) есть добавочные органы солевого обмена — надорбитальные солевые железы , выделяющие избыток солей в виде капелек секрета, стекающего из ноздрей .

Дыхательная система птиц также несёт признаки приспособления к полёту, во время которого организм нуждается в усиленном газообмене . Эта система органов у птиц считается одной из самых сложных среди всех групп животных . Дыхательные пути включают носовую и ротовую полости, верхнюю гортань, трахею, бронхи и сиринкс . От глотки отходит длинная трахея, делящаяся в грудной полости на два бронха. На месте бифуркации трахеи имеется расширение — сиринкс (нижняя гортань). Она представляет собой голосовой аппарат и наиболее развита у поющих и издающих громкие звуки птиц. Лёгкие птиц невелики по объёму, малоэластичны и прирастают к рёбрам и позвоночнику . Они имеют трубчатое строение и очень густую капиллярную сеть. С лёгкими связаны воздушные мешки — тонкостенные, легко растяжимые выросты вентральных ответвлений крупных бронхов, находящиеся среди внутренних органов, между мышцами и заходящие в полости костей . У большинства птиц 9 воздушных мешков . Они играют большую роль в дыхании во время полёта . Кроме того, они участвуют в терморегуляции, примыкая к крупным группам мышц и отводя лишнее тепло, а у водоплавающих птиц повышают плавучесть .
Дыхательная система устроена таким образом, что воздух проходит через лёгкие насквозь, причём в одном и том же направлении и на вдохе, и на выдохе . При вдохе только 25 % воздуха остаётся в лёгких, а 75 % поступает по бронхам в воздушные мешки . Этот воздух проходит через лёгкие на выдохе (т. н. двойное дыхание). Таким образом, лёгкие всегда омываются свежим воздухом, что улучшает газообмен . В состоянии покоя дыхание птицы осуществляется путём расширения и сжатия грудной клетки. При полёте, когда движущимся крыльям нужна твёрдая опора, грудная клетка птиц остаётся практически неподвижной и прохождение воздуха сквозь лёгкие обеспечивается расширением и сжатием воздушных мешков . Чем интенсивнее машущий полёт, тем интенсивнее и процесс дыхания. При подъёме крыльев воздушные мешки растягиваются, и воздух засасывается в них и в лёгкие. При опускании крыльев происходит выдох, и через лёгкие проходит воздух из воздушных мешков . Впрочем, полная синхронизация дыхания и движений крыльев достигается лишь при наиболее интенсивном дыхании .

Голосовой аппарат птиц имеет не одну гортань , а две — верхнюю ( larynx ) и нижнюю ( syrinx ). Главная роль в образовании звуков принадлежит нижней, наличие которой отличает птиц от других животных. Располагается она в месте разветвления трахеи на два главных бронха .
Нижняя гортань позволяет использовать трахею в качестве сильнейшего резонатора. У многих птиц последняя сильно увеличивается в длину и в диаметре, увеличиваются также и бронхи, каждый являющийся независимым источником звука. Птица способна значительно изменять форму сложной системы голосового аппарата при помощи движений тела и напряжения специальных мышц. Это позволяет ей управлять высотой и тембром голоса . Голосовой аппарат занимает значительную часть тела, что особенно характерно для маленьких птиц, у которых в процесс пения вовлекается весь организм .

Кровеносная система замкнутая, имеет два круга кровообращения. Сердце четырёхкамерное, состоит из двух предсердий и двух желудочков, с полным разделением артериальной и венозной крови . В отличие от млекопитающих, у птиц главной артерией является правая (а не левая ) дуга аорты , с которой начинается большой круг кровообращения . Проходящая через конечности кровь далее попадает в почечно-портальную систему, а затем в нижнюю полую вену . От дуги аорты отходят парные безымянные артерии, которые затем делятся на сонные артерии, мощные грудные и подключичные артерии, идущие к грудным мышцам и крыльям. От спинной аорты ответвляются артерии к туловищу и к ногам. Венозная система птиц в основном сходна с таковой у пресмыкающихся .
Частота сердечных сокращений у птиц высока, особенно у мелких видов. В полёте пульс заметно учащается по сравнению с покоем. Так, у серебристых чаек в покое частота сердечных сокращений достигает 250 в минуту, при беспокойстве возрастает до 450 уд/мин, а в планирующем полёте — 200—300 уд/мин . У воробья в покое пульс составляет около 460 уд/мин, а в полёте — около 1000 уд/мин .
Селезёнка у птиц относительно мала и обычно прилегает к правой стороне желудка. По форме, окраске и функциям она сходна с селезёнкой рептилий .
Лимфатическая система птиц характеризуется слабым развитием лимфатических сосудов. Они от всего тела в конечном счёте сливаются в два ствола: левый и правый грудные млечные протоки, которые соединяются между собой поперечными веточками и впадают в правую или левую краниальные полые вены. Лимфатических узлов у преобладающего большинства птиц нет; они отмечены только у гусеобразных и некоторых других .
Нервная система, в связи с приспособлением к полёту и обитанием в разнообразных условиях, развита у птиц значительно лучше, чем у пресмыкающихся . Головной мозг намного больше, чем у пресмыкающихся. Его масса составляет у бескилевых птиц и пингвинов 0,04—0,09 % от массы тела, а у остальных летающих птиц — от 0,2 до 5—8 % от массы тела .
Продолговатый мозг развит очень хорошо: здесь находятся центры, регулирующие дыхание, кровообращение, пищеварение и другие жизненно важные функции, которые у птиц осуществляются особенно интенсивно. Мозжечок , расположенный в задней части головного мозга, велик, что связано с полётом, требующим очень точно координированных движений и постоянного сохранения равновесия тела.

Средний мозг сдвинут на вентральную сторону, прикрыт сверху большими полушариями и виден только по бокам; как правило, хорошо развиты его зрительные доли. Промежуточный мозг мал, эпифиз слабо развит, гипофиз крупный.
Конечный мозг сильно развит, это самая большая часть головного мозга . Кора конечного мозга у птиц выражена лучше, чем у пресмыкающихся, но её слой довольно тонок. Хорошо развиты полосатые тела — главная часть переднего мозга, в которых расположены центры регуляции зрения, движения и т. д. Обонятельные доли малы и тесно примыкают к большим полушариям, поэтому способность птиц к восприятию запахов ограничена , хотя у киви и птиц- падальщиков обонятельные доли развиты гораздо лучше. Полушария передней части мозга гладкие, без извилин и по сравнению с млекопитающими относительно невелики. В их функцию входит управление поведением, ориентация в пространстве, употребление пищи, спаривание и способность строить гнёзда. По современным взглядам, значительную часть больших полушарий у птиц занимает сложно дифференцированный паллиум . Ранее большая его часть принималась за производные полосатых тел. У птиц паллиум содержит участки, гомологичные слоям неокортекса млекопитающих, но располагаются они не слоями, а образуют так называемые «ядра» .
Спинной мозг в связи с отсутствием хвоста относительно короткий. Он образует два утолщения: шейное и поясничное (крестцовое), от которых отходят нервы к передним и задним конечностям, образующие по пути мощное плечевое и тазовое нервные сплетения .
Хорошо развитый головной мозг позволяет птицам вырабатывать сложные формы поведения и приспосабливаться к самым разным ситуациям. Наиболее яркими проявлениями умственных способностей птиц считаются обучение певчих птиц пению, повторение человеческой речи попугаями , приёмы добывания корма у ряда видов и умение врановых решать сложные задачи, предлагаемые им в специальных экспериментах . Установлено, что у птиц способность к счёту развита лучше, чем у млекопитающих [ уточнить ] .

Например, в Англии синицы, обитавшие около загородных домов, научились снимать алюминиевые крышки с выставленных бутылок с молоком либо продырявливать их и склёвывать сливки . Описано для птиц и использование инструментов. Дятловый древесный вьюрок , обитающий на Галапагосских островах , держа в клюве крупную иглу кактуса или сухую веточку, ковыряет ею в трещинах коры для извлечения насекомых из глубоких расщелин в древесине. Перелетая с дерева на дерево, вьюрок часто переносит колючку с собой . Новокаледонский ворон — один из немногих видов, способных не только использовать, но и изготавливать орудия труда , с помощью которых птица достаёт из-под коры личинок насекомых .
Некоторые птицы обладают особыми методиками привлечения добычи. Например, зелёные кваквы бросают в водоём лист, веточку или перо и следуют за ними по течению, ожидая, пока «приманкой» не заинтересуется рыба .
В Африке обыкновенный стервятник , найдя страусиное яйцо, скорлупу которого он не может разбить клювом, разыскивает камень (иногда массой до 0,5 кг при массе птицы 2—2,5 кг) — и, многократно бросая его клювом на яйцо, раскалывает скорлупу . Иногда вороны и крупные чайки, схватив твёрдую добычу (орехи, моллюски, крабы и др.), взлетают и затем бросают добычу на землю. Приём повторяется многократно, пока орех, раковина или панцирь не расколется. Так же поступают и некоторые хищные птицы с черепахами (стервятники) или с крупными костями ( бородач ). Различные виды дятлов используют щели в стволах деревьев для закрепления раздалбливаемых ими шишек. Певчий дрозд использует камни в качестве наковальни для того, чтобы разбивать раковины моллюсков .
Обыкновенная сорока — единственная птица, прошедшая зеркальный тест . Попугаи-жако , галка и новокаледонский ворон не прошли этот тест, хотя временами демонстрировали некоторое поведение, связанное с зеркалом [ какое? ] .

Зрение — самое важное для птиц чувство, а глаза — основные органы ориентирования. Многие птицы хорошо видят вдаль ( сокол-сапсан способен увидеть небольшую птицу на расстоянии более 1 км). У некоторых видов поле зрения достигает почти 360°. Зрение у птиц заметно острее, нежели у других позвоночных — это объясняют значительно большим количеством светочувствительных клеток в сетчатке глаза . Глаза могут двигаться независимо друг от друга, но у большинства видов они малоподвижны (у некоторых, например сов, вовсе неподвижны), поэтому птицам приходится часто двигать головой, чтобы хорошо рассмотреть окружающую среду . Как правило, зрение монокулярное , зато с очень большим полем зрения — у обыкновенного и американского вальдшнепов его угол может достигать 360° в горизонтальной плоскости и 180° в вертикальной, охватывая, таким образом, всю сферу . Область, доступная обоим глазам, у большинства птиц невелика. Максимальной ширины (около 50°) она достигает у совообразных , и у них, как и некоторых других хищных птиц, показано наличие стереоскопического зрения . Глаза ныряющих птиц имеют особенно гибкие хрусталики , приспособленные к зрению как в воздухе, так и под водой . Все птицы обладают цветным зрением. Важная особенность зрения птиц — тетрахроматия : в сетчатке глаза есть пигменты с максимумом поглощения в красной, зелёной, синей и ближней ультрафиолетовой областях . Кроме того, криптохром , служащий для восприятия синего цвета, воспринимает магнитное поле Земли , позволяя его буквально видеть; молекула быстро переключается между сигнальным и неактивным положением благодаря присутствию супероксида , содержащегося в чуть более высокой концентрации, чем у млекопитающих .

Наиболее подвижным веком является, как правило, нижнее. Кроме наружных век, у птиц, подобно рептилиям, есть «третье веко» — мигательная перепонка , которая располагается в переднем углу глаза и двигается по горизонтали . У многих водных птиц перепонка полностью закрывает глаза и под водой выполняет функцию защитных очков .
Органы обоняния и вкуса сравнительно слабо развиты. Вкусовые органы птиц представлены вкусовыми почками , располагающимися на мягком нёбе и у основания языка. У подавляющего большинства видов плохо развито чувство обоняния, они практически не различают запахи . Однако у немногих наземных птиц ( киви , кулики) и у некоторых других групп (трубконосые, некоторые дневные хищники, американские грифы ) обоняние довольно развито .

Ухо птиц по строению сходно с ухом рептилий; оно значительно проще, чем ухо млекопитающих, но не менее эффективно . Ушных раковин нет, но у некоторых птиц ухо обрамлено складкой кожи или перьями, образующими их подобие (развиты у ушастых сов , филинов и со́вок ) . Барабанная перепонка лежит в небольшом углублении, прикрытом перьями. Среднее ухо содержит одну косточку , которая передаёт звуковые волны от барабанной перепонки в улитку внутреннего уха . Улитка не закручена и выглядит как продолговатый вырост круглого мешочка . Прекрасно развито у птиц восприятие изменений положения тела, благодаря хорошему развитию соответствующих частей лабиринта.
Самый известный пример птиц, способных к эхолокации , — южноамериканские гуахаро . Ею владеют и саланганы ( и Aerodramus ), гнездящиеся на отвесных скалах и в пещерах Юго-Восточной Азии и островах Борнео и Шри-Ланка . Эхолокация помогает этим птицам ориентироваться в темноте .
Органы осязания рассеяны в разных местах поверхности кожных покровов и представлены главным образом тремя типами образований: тельцами Грандри , Меркеля и Гербста .
У птиц доказано наличие как минимум двух независимых систем магниторецепции . Одна система находится в сетчатке глаза и, вероятно, основана на фотозависимых химических реакциях на основе криптохрома . Зрительные магниторецепторы обеспечивают восприятие компасной информации, требующейся для ориентации птиц при миграциях. В основе второй системы, вероятно, лежат магнитные свойства соединений железа (биогенного магнетита ) . Она предположительно расположена в надклювье и участвует в репрезентации пространства, обеспечивая аналог карты. Некоторые авторы предполагают наличие третьей системы магниторецепции, основанной на вестибулярном аппарате . Не исключается, что магниторецепция на основе оксидов железа происходит в лагене — структуре, гомологичной улитке внутреннего уха млекопитающих, а информация от них обрабатывается в вестибулярных ядрах головного мозга .
Птицы являются раздельнополыми. Их пол определяется половыми хромосомами Z и W , а не X и Y, как у млекопитающих. Самцы птиц несут две Z-хромосомы (ZZ), а самки — W-хромосому и Z-хромосому (WZ) .
Органы размножения самца представлены парными семенниками бобовидной формы, лежащими в брюшной полости . От них отходят семяпроводы , открывающиеся в клоаку . Размеры семенников изменяются в течение года: в период размножения их вес может увеличиваться в 250—300 раз. У большинства птиц внутреннее оплодотворение обеспечивается тем, что отверстие клоаки самца прижимается к клоаке самки, в которую впрыскивается порция спермы. Наружные половые органы , как правило, отсутствуют (исключение — аргентинская савка ). У некоторых птиц ( страусообразные , тинаму , гусеобразные и краксы из курообразных) от стенки клоаки обособляется непарный совокупительный орган, имеющий фиброзные тела; при совокуплении он выдвигается и вводится в клоаку самки .
Органы размножения самок представлены обычно только одним, левым, яичником , лежащим вблизи почки. У самок некоторых попугаев и хищных птиц иногда бывает и правый яичник. Яичник имеет зернистое строение. Формирование фолликулов заканчивается в первые месяцы постэмбриональной жизни (у галки образуется около 26 000 фолликулов). Ежегодно к началу размножения начинается рост немногих фолликулов (в 5—10 раз больше нормального числа откладываемых яиц), и яичник приобретает гроздевидную форму. Яйцевод представляет собой длинную трубку, открывающуюся воронкой в полость тела около яичника; другой конец яйцевода открывается в клоаку . Вышедшее из яичника яйцо попадает в непарный яйцевод, в верхней части которого и происходит оплодотворение . Пройдя по яйцеводу, яйцо приобретает белковую оболочку, а попав в более широкую матку, покрывается скорлупой. Через влагалище — конечный отдел половых путей самки — яйцо попадает в клоаку, а оттуда уже выводится наружу .


Половой диморфизм у птиц широко распространён и часто резко выражен . Он проявляется обычно по достижении птицами половой зрелости, и лишь изредка — в первом весеннем наряде. Особенно ярко половой диморфизм выражен в брачный период . Пингвины, веслоногие, буревестники, поганки, гагары, вертишейки, стрижи, многие зимородки и щурки не имеют полового диморфизма . Самцы и самки мелких воробьиных, большинства хищных птиц, сов, чаек, куликов, чистиков, пастушков и др. отличаются только размерами (обычно самцы крупнее самок). Широко распространены различия в окраске самцов и самок. Обычно половой диморфизм резче выражен и окраска самцов более ярка у тех видов, самцы которых не принимает непосредственного участия в заботе о потомстве. У многих птиц диморфизм окраски имеет место только в период размножения. Половой диморфизм в окраске часто выражается в цвете не только самого оперения, но и других частей тела птицы (клюв, радужина, голые участки кожи) .
Половые различия также выражаются в наличии выростов и придатков кожи на голове (у куриных), в форме отдельных частей тела, в развитии отдельных перьев (хохолки, длинные кроющие перья надхвостья павлина, перья на крыльях и хвосте райских птиц , длинные рулевые у фазановых и т. д.), в пропорциях и размере отдельных частей тела, в строении внутренних органов (горловой мешок фрегатов и т.п) . Половой диморфизм (в том числе и брачный наряд ) играет важную роль в нормальном течении полового цикла и прежде всего облегчает опознание и нахождение брачного партнёра .

Жизнь птиц слагается из многократно повторяющихся периодических явлений, среди которых есть суточные и годичные. Суточные представляют собой чередование периодов активности и отдыха. Годичные же гораздо сложнее, отражают приспособление организма к циклически меняющимся условиях. Таковы явления постройки гнёзд, линьки и миграции .
Спектр питания птиц широк и включает разнообразные растительные и животные корма . От характера питания в значительной мере зависит и географическое распространение птиц . Основную роль в поисках пищи играет зрение, отчасти слух (лесные и ночные виды), порой осязание (кулики, утиные, фламинго) . Преобладающее большинство птиц добывают корм на земле либо на растениях, меньшинство — в воде или воздухе (на лету) .
По разнообразию используемых кормов птиц делят на три группы . Полифаги (всеядные) питаются разнообразными растительными и животными кормами. К этой группе можно отнести представителей примерно трети всех семейств, причём в пределах каждого семейства всеядность сильнее выражена у более крупных видов. Примером могут быть крупные врановые , крупные чайки, журавли и др .


Противоположной группой являются стенофаги — птицы, потребляющие однородные корма и использующие однообразные приёмы добычи корма. Стенофагия встречается среди птиц редко. Она наблюдается у стрижей и многих козодоев , питающихся только летающими насекомыми, и ласточек . К этой же группе относятся типичные падальщики — грифы и марабу , а также виды, питающиеся преимущественно крупной рыбой — пеликаны , скопа , бакланы и др . Африканский пальмовый гриф питается плодами нескольких видов пальм ; коршун-слизнеед поедает лишь моллюсков . Нектаром цветов ограниченного числа растений питаются некоторые виды мелких колибри , медососов , нектарниц и гавайских цветочниц . Клесты питаются семенами хвойных деревьев .

Большинство птиц относится к промежуточной группе и питаются довольно широким спектром кормов. Таковы многие воробьинообразные , потребляющие насекомых и семена, курообразные , туканы и птицы-носороги, питающиеся зелёными частями растений, семенами, ягодами и беспозвоночными .
По составу корма птиц также разделяют на ряд экологических групп . Виды, питающиеся преимущественно растительной пищей — фитофаги . Почти исключительно листьями питается гоацин ; гуси, лебеди, некоторые утки, лысухи питаются главным образом разнообразной прибрежной и водной растительностью, попутно поедая водных животных. Преимущественно семенами питаются многие воробьинообразные — ткачиковые , вьюрковые , жаворонковые и др. Семенами, ягодами и плодами питаются многие попугаи, голуби, турако , птицы-мыши , многие трогоновые , райские птицы . Однако большинство этих птиц выкармливает птенцов преимущественно животными кормами .
Виды, питающиеся преимущественно животной пищей, называют зоофагами , хотя многие из них в небольшой степени поедают и растительные корма. Почти треть всех семейств птиц исключительно или преимущественно насекомоядны ( энтомофаги ). Однако насекомыми в той или иной степени питаются почти все птицы. Многие водные и околоводные птицы питаются преимущественно рыбой , попутно поедая и водных беспозвоночных .
Многие хищные птицы и совы питаются мелкими грызунами . Немногие хищные, например сапсан , ястребы и чеглок , болотный лунь и некоторые другие питаются преимущественно другими птицами. Небольшое число видов птиц питается моллюсками (например, арама ). К герпетофагам, питающимся амфибиями и рептилиями, относятся орёл-змееяд , птица-секретарь , кариамы , некоторые крупные зимородки ( кукабары ) и др. Некоторые чайки и качурки относятся к копрофагам — могут поедать помёт китообразных и ластоногих . Гарпии-обезьяноеды ловят обезьян и ленивцев , красногорлый сокол и широкоротый коршун охотятся преимущественно на летучих мышей . Малайский орёл-яйцеед таскает яйца из гнёзд других птиц .
В связи с сезонностью появления разных типов кормов у многих видов птиц наблюдается сезонное изменение рациона. Это особенно характерно для зимующих и перелётных птиц в северном полушарии. У многих птиц рацион различен в разных частях ареала, поскольку доступность кормов зависит не только от времени, но и от региона .
Птицы некоторых видов запасают корм. Это может касаться даже животной пищи: воробьиный и мохноногий сычи осенью складывают в дупла пойманных зверьков и птиц. Кедровка прячет в дуплах и других местах кедровые орешки . Сойки прячут жёлуди среди опавших листьев, в пнях, дуплах и т. п. Желудёвый дятел из США выдалбливает в стволах деревьев отверстия, в которые забивает собираемые им жёлуди. В одной жёлтой сосне было обнаружено до 60 тысяч спрятанных им желудей . Подобные запасы пищи используются птицами в зимнее время .

Мелкие птицы нуждаются в относительно большем количестве пищи, чем крупные. Масса пищи, потребляемой мелкими воробьиными в сутки, составляет 50-80 % их массы. Хищные птицы массой около 1,5 кг нуждаются в 150—200 г мяса, однако поймав крупную добычу, могут съесть до 800—1000 г. Мелкие птицы погибают без пищи уже через 15—30 часов, голуби — через 7—9 дней, крупные орлы и совы могут голодать до месяца .
Потребность птиц в воде различна. Кожное испарение птиц незначительно, кроме того, вода из мочи всасывается обратно, когда моча находится в верхнем отделе клоаки . Многие пустынные и степные виды могут обходиться без воды месяцами (например джек , пустынная славка ) . Однако большинству других видов вода необходима каждый день.
Морские птицы могут пить морскую воду благодаря специальным железам , выводящим избытки соли . Большинство птиц зачерпывают воду клювами и поднимают голову, чтобы вода текла в горло. Некоторые виды, например голуби, рябки, птицы-мыши , трёхперстки , дрофиные и другие способны втягивать воду, не запрокидывая голову . Многие виды рябков ( чернобрюхий рябок , южноафриканский рябок ) приносят воду своим птенцам не только в клюве, но и на перьях брюшка, способных удерживать воду благодаря особому строению .
Звуковые сигналы имеют в жизни птиц исключительно важное значение. Ими обеспечиваются охрана кормовой и гнездовой территории от вторжения чужаков, привлечение самки для выведения потомства, предупреждение сородичей и птенцов о грозящей опасности. В языке птиц насчитываются десятки звуковых сигналов (бедствия, предостережения, пищевые, ухаживания, спаривания, агрессивные, стайные, гнездовые и так далее) .

Птицы могут издавать звуки не только голосовым аппаратом, но и с помощью клюва, лап, крыльев, хвоста или посторонних предметов (например, дятлы) .
Песню птицы принято делить на отдельные элементы, называемые нотами или слогами. Большинство песен состоит из 1—2 слогов, однако бывает и больше (13—24 слога у пеночки-теньковки ). Более сложные части, состоящие из нескольких слогов, называются мотивами, фразами или коленами (как у соловья) . Одна и та же песня в период размножения может служить сигналом привлечения самки, а в период высиживания птенцов — сигналом занятости территории .
В вокализации выделяют песни и позывы , отличия между которыми в некоторой мере условны. Песни более длинные, сложные, разнообразные и связаны в основном с брачным и территориальным поведением, тогда как позывы выполняют и много других функций (предупреждения, удержания стаи вместе и т. д.) . Пение наиболее развито у птиц отряда воробьинообразных. Обычно поют самцы , а не самки , хотя встречаются исключения. Чаще всего птицы поют, сидя на каком-либо субстрате, хотя некоторые виды способны петь в полёте . Птицы некоторых групп молчаливы, издают лишь механические звуки, например аист только щёлкает клювом . У некоторых манакинов самцы развили несколько механизмов образования таких звуков, включая стрекотание подобно насекомым , но с помощью перьев .
Целенаправленно издаваемые звуки, образованные не сиринксом , а другими частями тела, называются механическими , или инструментальными звуками , а также сонациями .
Многие птицы способны перенимать песни других видов и имитировать голос человека (включая индивидуальные особенности его тембра, как это делают скворцы, попугаи, вороны и др.) . Существуют и случаи присвоения голосовых сигналов, а не пения птиц. Например, панамский черноспинный лесной сокол способен имитировать призывы своих жертв, а потом ловит птиц, которые прилетели в ответ на призыв . Лирохвосты обладают феноменальной способностью имитировать всевозможные звуки, начиная от голосов птиц, других животных, человека, и заканчивая различными техногенными звуками, включая игру на флейте и даже звуки автосигнализации и бензопилы .


Полёт — основной способ передвижения большинства видов птиц, помогающий им искать пищу, мигрировать и спасаться от хищников . Это наиболее специфическая форма передвижения, определившая основные черты организации всего класса . Способность птиц к полёту вызвала ряд адаптаций для обеспечения полёта, взлёта и посадки, ориентирования в пространстве и навигации . Возможность полёта ограничивает максимальную массу летающих птиц. Чем крупнее и тяжелее птица, тем больше энергии она тратит на взмахи крыльями; начиная с определённой массы для полёта требуется больше энергии, чем птица может произвести . Поэтому для активного машущего полёта птица должна весить не слишком много. Масса современных летающих птиц обычно не превышает 2 кг, изредка достигает 12—15 кг :114 и максимальна (18-19 кг) у дрофиных .
Лишь немногие птицы (около 40 современных видов ) в ходе эволюции потеряли способность к полёту . Некоторые из этих птиц слишком велики для полёта, а некоторые обитают на изолированных островах, где нет наземных хищников . Потеря способности к полёту у бескилевых вызвала редукцию киля грудины :161 . К нелетающим птицам не относят мало летающих ( курообразные и др.), так как они не утратили способность летать, хотя и не совершают дальних перелётов.
Полёт птиц принято разделять на два основных типа : активный (машущий) и пассивный (парящий). Птицы обычно используют не один тип полёта, а комбинируют их. За взмахами крыльев следуют фазы, когда крыло не совершает движений: это скользящий полёт, или парение. Такой полёт характерен преимущественно для птиц средних и крупных размеров, с достаточной массой тела . Парящим называется полёт без активных затрат энергии со стороны птицы. Различают парение динамическое и статическое. В основе статического лежит использование потоков обтекания или воздушных термических потоков. Ещё один тип парения связан с использованием птицами термиков — потоков восходящего воздуха. Такое поведение характерно для многих хищных птиц , в частности грифов , коршунов , канюков . Этим методом также пользуются и аисты , пеликаны и другие птицы, которые не являются хищными.


Зависание на месте ( англ. hovering , трепещущий полёт ) — сложная для птиц задача. Большинство птиц или совсем неспособны к нему или способны лишь на протяжении очень короткого времени. Фактически единственная группа птиц, приспособленных к этому, — колибри . Некоторые птицы способны зависать относительно поверхности за счёт полёта против ветра со скоростью ветра ( англ. windhovering ). Такое поведение характерно для многих хищных и рыбоядных птиц: буревестников, скоп, крачек , поморниковых , некоторых зимородков .
По ветвям, земле и воде птицы передвигаются с помощью задних конечностей. Только у стрижей все четыре пальца лапы направлены вперёд, заканчиваются острыми изогнутыми коготками и служат только для цепляния и опоры; передвигаться по земле и ветвям они не могут . Бедренные кости птиц коротки, малоподвижны и почти не смещаются из горизонтального положения (благодаря чему помогают поддерживать и, возможно, вентилировать брюшные воздушные мешки). Это одна из основных характеристик птиц; у остальных животных, передвигающихся по земле, тазобедренный сустав намного подвижнее .
Большинство мелких древесно-кустарниковых птиц передвигаются по земле и в ветвях прыжками. Птицы средних размеров (голуби, вороны и др.) и некоторые мелкие птицы ( трясогузки и др.) по земле ходят и бегают, попеременно переставляя ноги. Ряд видов чередуют ходьбу и прыжки. У птиц наземного образа жизни ( фазановые , тинаму , дрофы , рябки и др.) крепкие пальцы относительно короткие и толстые, задний палец часто мал и не служит для опоры. По земле они ходят и бегают, попеременно переставляя ноги; на ветви садятся лишь немногие виды .

Любая птица в воде не тонет и при необходимости плывёт . Могут присаживаться на воду дрозды и голуби , довольно часто плавают пастушки и многие кулики , иногда садятся на воду и плавают цапли и т. д. Но у птиц, постоянно связанных с водой, развиваются приспособления, облегчающие плавание, а у многих видов — и ныряние . Низкий удельный вес птиц, облегчая плавание, затрудняет ныряние. Некоторые птицы при ловле добычи способны нырять с пикирования: птица полускладывает крылья и резко пикирует, погружаясь в воду на небольшую глубину по инерции. К такому нырянию прибегают птицы, которые могут плавать — трубконосые , многие пеликанообразные . Ныряют также птицы, обычно не садящиеся на воду — скопа , орланы, рыбный филин , многие зимородки. По-настоящему погружаться на большую глубину и активно передвигаться в толще воды может относительно небольшое количество видов хорошо плавающих птиц. Обычно птицы могут нырять на глубину нескольких метров. Некоторые нырковые утки (гаги, морянки и др.), кайры , гагары , поганки , пингвины ныряют иногда на глубину 50—60 м и крайне редко — несколько глубже . Погружения птиц обычно длятся не более 1—1,5 минут , но у пингвинов и гагар достигают 10—15 минут и более. Рекорд, зафиксированный для птиц — погружение императорского пингвина на глубину 564 м, продолжавшееся 21,8 минуты . Из воробьиных птиц ныряет оляпка . Опустившись на воду, она держит крылья так, что течением её прижимает ко дну, и она бежит, переворачивая камни и склёвывая добычу под ними. К такому своеобразному нырянию в быстро текущую воду и беганью по дну способны также вилохвостки и водяные печники .

В неволе многие воробьиные птицы доживают до 20—25 лет, цапли, аисты, некоторые куриные, лебеди, гуси — до 30—35 лет, пеликаны, журавли, серебристая чайка, крупные хищники — до 40—50 лет, крупные попугаи, филины, вороны — до 70 лет. В естественных условиях обитания максимальная продолжительность жизни птиц значительно меньше, и её достигают лишь отдельные особи. Средняя продолжительность жизни в диких условиях, вероятно, меньше максимальной в 5—10 раз .
Наступление половозрелости у птиц зависит от их величины: чем крупнее птица, тем позднее она приступает к размножению. Самки мелких ткачиков способны к оплодотворению уже в возрасте 6 месяцев. Большинство воробьиных становятся половозрелыми в 8—12 месяцев; вороны, мелкие чайки и крачки, мелкие дневные хищники и утки — на втором году жизни; крупные чайки, веслоногие и гусиные, орлы, гагары — к концу третьего года, страус — на четвёртом году жизни .
В жизни птиц процесс размножения вызывает развитие различных по своему биологическому смыслу приспособлений, обеспечивающих воспитание и выживание потомства. Период размножения у птиц выпадает так, что появление птенцов приходится на время года с наибольшим количеством доступного корма (для данного вида на определённой территории) . В тропиках, в условиях мало или не изменяющейся продолжительности светового дня, время размножения определяется в основном уровнем влажности — большинство птиц в этом регионе гнездится в сезон дождей .
Все птицы яйцекладущие . Их яйцеклетка увеличивается и превращается в большое яйцо , которое содержит запас питательных веществ, необходимый для развития зародыша . Оплодотворение происходит внутри. После яйцекладки для продолжения развития зародышу необходимо тепло, поэтому родители согревают его теплом своего тела на протяжении нескольких недель или даже месяцев .
Примерно у 90 % видов птиц отмечается моногамия — пары формируются на один или реже на несколько сезонов размножения (преимущественно певчие птицы), у некоторых групп птиц — на всю жизнь (крупные хищники, совы, цапли, аисты и др.). Существуют и такие виды, которые пар не образуют вовсе и у которых всю заботу о потомстве осуществляют особи только одного пола, чаще всего самки. Это встречается у большинства куриных, многих уток, всех райских и беседковых птиц. У куликов-плавунчиков, трёхперсток, тинаму, яканы, африканского страуса о выводке заботится самец.

Полигамия встречается у птиц в виде полиандрии (несколько самцов и одна самка) и полигинии (несколько самок на одного самца). Первая форма отмечается у плосконосых плавунчиков , трёхперсток, якан, некоторых тинаму и др.; вторая — у дроздов , крапивников, некоторых славок , луговых трупиалов и др. При полиандрии самка откладывает несколько кладок яиц, которые насиживают исключительно самцы, которые также водят и выводки. При полигинии строят гнёзда и насиживают кладку только самки .
У многих моногамных видов и самец, и самка принимают более или менее одинаковое участие в постройке гнезда, насиживании кладки и выкармливании птенцов (трубконосые, голенастые, чайки, кроншнепы, городские ласточки и многие другие), у других — самка строит гнездо и насиживает кладку, а самец занимается охраной гнездового участка, иногда приносит строительный материал и кормит самку или на протяжении коротких периодов насиживает кладку. Обычно выкармливают птенцов оба партнёра (пеликаны, журавли, многие воробьиные). У некоторых типичных моногамных птиц изредка встречается бигамия: на участке одного самца поселяются две самки. В ряде случаев самец принимает участие в насиживании и выкармливании птенцов обеих самок, в других — вторая самка самостоятельно занимается выращиванием потомства .
У полигамных птиц пары не образуются даже на короткий период: в период яйцекладки самки могут спариваться с различными самцами; процессом постройки гнезда, насиживанием и заботой о птенцах занимается самка . К полигамным птицам относится большинство тетеревиных (рябчик и белые куропатки — моногамны) и других курообразных, некоторые дрофы, ряд куликов (турухтан и др.), почти все виды колибри, некоторые каменные петушки, лирохвосты , большинство райских птиц и многие другие .
С наступлением периода размножения происходит рассеивание стай, занятие птицами отдельных гнездовых территорий, образование пар у моногамных видов и закрепление за каждой парой своего гнездового участка, на котором находится гнездо и на котором гнездящиеся птицы будут придерживаться оседлого образа жизни на протяжении всего периода размножения . Образование пар у мигрирующих моногамных видов происходит на зимовках либо на весеннем пролёте. Поэтому к местам гнездования они прилетают уже сформировавшимися парами. У большинства же птиц пары обычно формируются на местах гнездования. У многих голенастых, воробьиных и других самцы прилетают раньше самок, занимая подходящие гнездовые участки, охраняя их и издавая брачные крики, совершая своеобразные полёты. Прилетающих позднее самок привлекает брачное поведение самцов, и они оседают на приглянувшихся участках .
| Размножение и развитие птиц на примере плачущей горлицы | ||||
|---|---|---|---|---|

|

|

|

|

|
| Кладка яиц в гнезде | Насиживание | Птенцы | Молодая птица | Взрослая птица |


Начало периода размножения у птиц проявляется своеобразным поведением, которое называют токовым. Во время токования самец, — а у многих видов и присоединившаяся к нему самка, — принимает различные необычные позы (распускание перьев, пряжки, бег с распущенными крыльями, вытягивание шеи и т. п.), совершает характерные движения («танцы» журавлей , «танцы» райских птиц, гонки по воде у лысух, чомг , уток и т. д.), интенсивно поёт либо издаёт громкие звуки и т. п. Порой такое поведение проявляется в своеобразном полёте (хищники, кулики, жаворонковые и мн. др.), в драках между самцами (тетерев, глухарь и др.) . У многих видов птиц отмечаются токовые полёты (белая куропатка, вальдшнеп и многие кулики, колибри и др.) — сложные передвижения в воздухе, сопровождаемые громкими звуковыми сигналами или пением. Бекас издаёт громкий блеющий звук путём вибрации узких крайних рулевых перьев. Сложные брачные ритуалы существуют у шалашников из Австралии и Новой Гвинеи , строящих в период размножения своеобразные шалаши, украшенные цветками, камешками, раковинами моллюсков, яркими плодами либо предметами одного цвета .
У некоторых полигамных видов птиц отмечается групповой ток самцов ( райские птицы , манакиновые и др.), часто сопровождающийся драками самцов (глухари, тетерева, турухтаны , скальные петушки и многие другие) .
Токование начинается обычно задолго до начала периода размножения и заканчивается только с вылуплением птенцов, либо же позже. Биологическое значение токового поведения заключается в облегчении встречи птиц одного вида для спаривания, в стимуляции развития половых продуктов самок и синхронизации половых циклов обоих полов .

Гнёзда птиц облегчают согревание яиц, обеспечивают защиту кладки, насиживающей птицы и птенцов от неблагоприятных погодных условий и хищников. Постройка гнёзд может быть чрезвычайно сложной. Птицы устраивают гнездо, чтобы отложить в нём яйца и вырастить птенцов. Однако существуют птицы, не устраивающие гнёзда, а другие не воспитывают в нём птенцов, так как те оставляют его сразу после вылупления из яиц. Большинство птиц гнездится в течение года один раз, многие по два и более. Перелётные птицы на зимовках не гнездятся .
Гнёзда птиц очень разнообразны по расположению, форме и применяемым для их постройки строительным материалам . Самый простой способ гнездования наблюдается у козодоев, которые откладывают яйца прямо на землю, даже не делая ямки. Гагарки и другие мелкие чистиковые гнездятся в расщелинах скал, не сооружая специальных гнёзд. Целый ряд птиц — речные и малые крачки, некоторые кулики — роют для гнезда ямку, но ничем её не вымащивают; у других куликов, куриных, дроф, журавлей и многих других птиц такая ямка имеет более или менее выраженную выстилку. Воробьиные, которые гнездятся на земле, вьют в ямке настоящее гнездо, а пеночка-трещотка к тому же делает своеобразную «крышу» над гнездом, которое благодаря этому имеет боковой вход. На наземные гнёзда похожи по строению плавающие гнёзда таких птиц, как поганки , гагары и болотные крачки. Эти гнёзда плавают по достаточно глубокой воде среди растительности и выдерживают вес кладки яиц и насиживающей её птицы. Значительно сложнее строение гнёзд птиц, которые гнездятся на деревьях и кустарниках. Но и здесь встречаются все переходы от простой кучки веточек ( голубиные ) до самых сложных гнёзд, среди которых есть свитые гнёзда зяблика и зелёной пересмешки , шаровидные гнёзда с боковым ходом длиннохвостой синицы и крапивника, висячие гнёзда иволги и ремеза .
Одни из наиболее искусно сплетённых гнёзд сооружают многие ткачиковые . Например, обыкновенный общественный ткач создаёт групповые гнёзда, где под единой крышей находится до 200—400 индивидуальных гнездовых камер. Обычно опорой для них служат деревья, но в Южной Африке ткачики охотно используют столбы линий электропередач .
Многие птицы гнездятся в закрытых местах. В дуплах сооружают гнёзда многие воробьиные, удоды, ракшеобразные, дятлообразные, совы, попугаи, некоторые голуби и др. Дятлы сами выдалбливают дупла для гнёзд, другие птицы используют природные или оставленные дятлами дупла. Многие птицы гнездятся в расщелинах скал или в домах (стрижи и др.). Ласточки, многие стрижи лепят гнёзда из смоченной почвы, а саланганы — из комочков быстро затвердевающей на воздухе слюны. Печниковые из Южной Америки устраивают из глины огромные шаровидные гнёзда на корнях мангровых деревьев. Некоторые птицы — береговые ласточки , зимородки, щурки — роют в обрывах с мягким грунтом норы с гнездовой камерой в конце хода .
Некоторые сорные куры не высиживают яйца, а закапывают их в землю или кучу гумуса, где они развиваются за счёт солнечного тепла или тепла, возникающего при гниении растений. Гнездо функционирует как инкубатор 6—9 месяцев. Некоторые виды сорных кур, живущие около действующих вулканов, закапывают яйца в тёплую почву на их склонах .
У многих птиц-носорогов после того, как самка отложит яйца, самец заделывает отверстие дупла с гнездом глиной, оставляя лишь узкую щель, через которую проходит клюв «пленницы». Самка насиживает яйца и продолжает оставаться в дупле несколько недель после вылупления птенцов .
У преобладающего большинства видов птиц развит гнездовой консерватизм: после успешного гнездования птицы на следующий год возвращаются на свой гнездовой участок, где восстанавливают своё старое гнездо или строят новое где-то поблизости :319 .
Многие птицы гнездятся колониально .





Самки некоторых птиц подкладывают яйца в гнёзда птиц другого, а иногда и своего вида, которые насиживают эти яйца и выкармливают птенцов. Это позволяет птице-паразиту тратить больше времени на поиски еды и размножение. Облигатные гнездовые паразиты никогда не строят гнёзд, не насиживают кладку и не кормят птенцов. Это отмечено у птиц 5 отрядов: у 50—80 видов кукушковых , всех 6 видов воловьих птиц , не менее 6 — медоуказчиковых , не менее 3 — вдовушковых , а также у черноголовой утки . У самых специализированных паразитов птенец избавляется от яиц и/или птенцов хозяина, который затем выкармливает только его.
Самки 32 видов гусеобразных подкладывают яйца в чужие гнёзда (своего или близких видов) лишь иногда — при нехватке подходящих мест для гнездования. Изредка это наблюдается и у некоторых чаек и куликов .

Питание зародыша обеспечивает желток . Существует два типа желтка — белый и жёлтый, которые организованы чередующимися концентрическими слоями. Желток заключён в вителлиновую мембрану и окружён белком. Содержимое яйца окружено двумя подскорлупковыми оболочками, внутренней и наружной. Снаружи яйцо покрыто скорлупой, состоящей преимущественно из карбоната кальция . После откладывания яйца на его тупом конце постепенно образуется воздушная камера .
Окраска яиц соответствует накоплению пигментов в разных слоях скорлупы . По окраске яйца разделяют на три группы: неокрашенные, одноцветно окрашенные и пёстрые. Первично скорлупа была неокрашенной, и это состояние сохранилось у ряда современных видов, преимущественно гнездящихся в дуплах, норах и других закрытых местах. Одноцветные яйца могут быть жёлтыми, чёрными (у ряда воробьиных), но чаще — различных оттенков голубоватого и зеленоватого цвета. Пятнистая окраска наиболее распространена и встречается у открыто гнездящихся птиц; она выполняет функцию маскировки .
Форма яиц разнообразна. Принято выделять пять типов форм яиц: равномерно эллиптические по обоим концам (козодои, колибри, др.); равномерно эллиптические с двумя заострёнными концами (казуары, нанду;) почти круглые (совы, кроме сипухи, щурки, зимородки); эллиптические с одним тупым концом и заострённым другим (большинство воробьиных, куриных, дневных хищников и др.); конические, грушевидные (кулики, чистики) . Относительные размеры яиц отличаются у разных видов. Мелкие птицы обычно откладывают относительно крупные яйца. Птенцовые птицы откладывают яйца меньших размеров, чем выводковые . Наибольшие размеры яйца были у вымерших эпиорнисов: масса 10 кг 460 г (вес скорлупы 3 кг 846 г), длина 340 мм, ширина 245 мм .
Количество яиц в кладке у разных птиц колеблется от 1 до 26. Так, буревестники, крупные хищники и пингвины, веслоногие и некоторые другие несут по одному яйцу, стрижи, козодои, голуби — по два, рябки и почти все чайки — по три. Кулики, кроме отдельных видов, откладывают по четыре яйца; воробьиные — чаще всего по пять-шесть, но синицы — по восемь и более (до 16). У уток чаще всего 6—10 яиц. Больше всего яиц в кладках перепелов и серых куропаток — до 22—26 . Самки многих видов птиц (утки, большинство воробьинообразных) откладывают яйца с интервалом в одни сутки. У многих курообразных, голенастых, чаек и гусей интервал составляет 24—48 ч, у журавлей, козодоев, кукушек — обычно 48 ч; у сов и хищных птиц — до 2—4 суток. Большинство птиц при гибели кладки откладывает повторную, часто — полную. У некоторых воробьиных и голубей в год бывает нормально две, а иногда — три и более кладок .
Раздел зоологии, посвящённый изучению яиц (преимущественно птичьих), называется оологией .

У птиц очень развита забота о потомстве, которая проявляется, кроме строительства гнезда и насиживания кладки, в выкармливании птенцов, в согревании и защите их от погодных условий, в очистке гнезда от экскрементов и более-менее активной защите от врага .
Обычно у полигамных птиц самец не участвует в заботе о потомстве, а у моногамных, напротив, всецело принимает в ней участие наряду с самкой .
Яйца чаще всего насиживаются самками, реже — обоими птицами из пары, совсем редко — только самцами. Насиживание начинается обычно после откладывания последнего яйца в кладке, но иногда ранее, в середине периода откладывания или уже после откладывания первого яйца (чайковые, пастушки и др.). Голенастые, хищные и совы, попугаи и ряд других птиц начинают насиживание сразу после откладки первого яйца . У мелких птиц период насиживания значительно короче, чем у крупных; среди последних некоторые насиживают дольше месяца. При насиживании у птиц на части живота и груди выпадает пух и образуется наседное пятно , которое обеспечивает более интенсивный нагрев яиц теплом тела .

В зависимости от продолжительности и сложности эмбрионального развития птицы подразделяются на две группы — выводковые и птенцовые .
Выводковые птицы (тинамуобразные, страусообразные, гусеобразные, курообразные, кроме гоацина, дрофы, многие кулики и др.) — птенцы которых вылупляются из яйца вполне сформированными, покрытыми пухом и способными отыскивать корм. Они тут же покидают гнездо, хотя ещё долгое время следуют за родителями, которые их защищают и помогают отыскивать корм .
Птенцовые птицы (веслоногие, дятлы, стрижи, попугаи, некоторые ракшеобразные и воробьинообразные) — птенцы которых вылупляются из яйца недосформированными, голыми, слепыми и беспомощными. Они долго остаются в гнезде. Родители не только защищают их, но и кормят из клюва. Такие птенцы вылетают из гнезда, когда они достигают или почти достигают размеров взрослых, а основные группы перьев почти завершают рост .
Между этими двумя типами развития есть и переходные — полувыводковые птицы (пингвины, трубконосые, большинство чистиковых) — у которых птенцы появляются отчасти сформированными, но долго остаются в гнезде и получают пищу от родителей .

Гибридизация встречается у птиц весьма часто — она отмечена между примерно 850 видами . Чаще всего это случается у близкородственных видов . Гибридизация у птиц наблюдается как в природе, так и при культивировании человеком (содержании в неволе). Наиболее известные примеры гибридов — межняк , лазоревка Плеске и мулард . Создание гибридов птиц человеком носит целенаправленный характер и направлено на выведение сельскохозяйственных птиц (мулард) либо ярко окрашенных особей птиц, содержащих в неволе с декоративной целью (гибриды попугаев, фазанов, певчих птиц). Гибриды птиц обычно способны давать плодовитое потомство.
В природе ухаживать за особями чужих видов пытаются преимущественно самцы, однако «чужие» самки их обычно отвергают — за исключением отдельных случаев. Если птиц-партнёров своего вида не хватает, особи любого пола и особенно находящиеся в низком социальном статусе или поздно прибывшие к местам гнездования существенно снижают селективность выбора партнёра. Шанс образования смешанных пар существенно выше у особей, не сумевших образовать нормальную пару в сроки, когда образуют пары большинство особей в данной популяции вида. Например, в зоне гибридизации серой и чёрной вороны особи с низким ранговым статусом могут выбирать для спаривания особей другого вида, если партнёры своего вида оказываются недоступными . Гибридизация также встречается в случаях, когда самка «ошибается» в выборе партнёра своего вида — случаи гибридизации у близкородственных видов певчих птиц. Ещё одна причина широкой межвидовой гибридизации — существенная схожесть брачных ритуалов и демонстраций у близких видов, или их полное совпадение.

Большинство птиц ведут дневной образ жизни. Строго ночных видов сравнительно мало; встречаются они преимущественно среди козодоев и совообразных. Исключительно ночной образ жизни ведут киви и попугай какапо .
Оцепенение для птиц не характерно, но существуют исключения. В оцепенение впадают на ночь колибри , обитающие в высокогорьях . У ласточек оцепенение может длиться несколько дней. Впадение в оцепенение, близкое к спячке, отмечается у белогорлого козодоя из США . Птенцы некоторых видов способны впадать в оцепенение в случае внезапного наступления холодов (птенцы воробьиных — до 2 дней, чёрного стрижа — до 5—8 дней).
Как и млекопитающие, птицы проявляют две фазы сна: медленный и быстрый (последняя фаза очень короткая — всего одна-две минуты ). У многих птиц фазы сна чередуются с моментами, когда птица быстро и кратковременно открывает глаза. Такой сон позволяет быть бдительной к опасностям . Медленный сон может протекать в одном или обоих полушариях головного мозга . Исследования подтвердили способность некоторых видов к глубокому сну только одного полушария мозга, а другое при этом остаётся активным . Висячие попугайчики часто отдыхают и спят на ветвях деревьев вниз головой .
Практически все виды птиц в период размножения проявляют хорошо выраженную территориальность — привязанность к своей гнездовой территории и её защиту от особей своего, а порой и других видов, включая хищников. Птицы добывают корм преимущественно на своей охраняемой территории (гнездовая и кормовая территории совпадают), но у некоторых видов кормовая намного обширнее :316 .
По характеру распределения пар на гнездовой территории и по удалённости гнёзд друг от друга птиц делят на одиночно гнездящихся и колониальных . У некоторых видов отмечается промежуточный тип гнездования — групповой (дрозды-рябинники, скворцы и др.) .


Одиночно гнездящиеся птицы располагают гнёзда на значительном расстоянии одно от другого. Соседние пары не связаны между собой и ведут относительно изолированный образ жизни. К этой группе принадлежит большинство птиц. У крупных хищников гнездовая территория занимает площадь нескольких квадратных километров, у мелких воробьиных птиц — 1000—8000 м 2 :316 .
Колониальные виды гнездятся большими скоплениями, гнёзда в которых располагаются близко друг от друга (от нескольких сантиметров до нескольких метров), образуя гнездовые колонии. Это свойственно видам, питающимся массовыми кормами и собирающим их на больших расстояниях от гнездовой колонии. Особенно много колониальных видов среди гнездящихся по побережьям морских птиц , образующих птичьи базары . Крупнейшая в мире колония бакланов и олушей, насчитывающая 10 млн особей, с плотностью до 4 гнёзд на 1 м², находится на побережье в Перу . Колониальность свойственна и ряду видов, населяющих берега крупных внутренних водоёмов ( фламинго , ряд бакланов, цапли, ибисы, чайки, крачки и др.). Реже колониальность встречается у наземных птиц: ряд ткачиков, грачи, береговые ласточки, розовые скворцы, гуахаро и др.
Многие колониальные птицы защищают свою колонию от хищников совместно. Например, так отгоняют хищников многие чайки и крачки. Около колоний птиц часто гнездятся и одиночно гнездящиеся виды. Многие виды птиц стремятся гнездиться и около одиночно гнездящихся видов, которые активно защищают свой гнездовой участок. Например, в стенках гнёзд орлов могут гнездиться воробьи; в близости от гнёзд хищных птиц гнездятся казарки, гуси, гаги, многие кулики. Это объясняется тем, что около гнезда ярко проявляется защитный рефлекс и подавляется рефлекс добывания пищи. Хищники не реагируют на поселившиеся рядом виды, на которых они активно охотятся вдали от своих гнёзд, и, активно защищая своё гнездо, защищают и своих «соседей» .

По характеру сезонных миграций все птицы делятся на перелётных , кочующих и оседлых . Первые ежегодно совершают перелёты в определённых направлениях с мест гнездования на зимовья и обратно, возвращаясь почти всегда в места гнездования в предыдущем году. Вторые в негнездовое время кочуют в ближайшие местообитания — такие кочёвки постоянных направлений не имеют. Третьи целый год оседло живут на одном участке. В одних частях ареала птицы определённого вида могут быть перелётными, а в других — вести кочевую или даже оседлую жизнь. Во время миграций на места зимовки и обратно к гнездовым областям перелётные птицы обычно придерживаются определённых направлений (миграционных путей), скапливаясь во время перелёта в экологически благоприятных местах .
Во многих крупных городах в умеренном климате сформировались популяции некоторых видов птиц, которые стали вести оседлый образ жизни, например кряквы . Все они гнездятся в самом городе или его окрестностях и отказались от сезонных миграций. Возникновение таких популяций связано с наличием доступного корма, в том числе с круглогодичной подкормкой птиц людьми, и отсутствием многих естественных врагов .
Среди сезонных миграций птиц, происходящих на протяжении года, выделяют следующие формы: послегнездовые кочёвки, осенне-зимние кочёвки, осенний перелёт, весенний перелёт .
Миграционные пути птиц состоят из множества отрезков, между которыми птицы отдыхают и ищут корм. Миграции изменчивы по срокам, а их траектории могут меняться из года в год и из сезона в сезон. Одна и та же особь в разные годы может мигрировать разными путями, а в отдельные годы не мигрировать вообще . Абсолютным чемпионом по дальности миграций считают полярную крачку , мигрирующую сезонно из Арктики в Антарктику, преодолевая за год до 70—90 тысяч км . Одну из самых длинных миграций в мире — до 26 тысяч км — совершает круглоносый плавунчик .
Большинство крупных птиц перелетают стаями, часто располагаясь геометрически правильным образом, таким как V-образный клин из 12—20 птиц. Такое расположение помогает им уменьшить энергетические затраты на перелёт. Незначительное число видов всегда летят поодиночке (некоторые дневные хищники, кукушки и др.).

Основной фактор, определяющий сроки прилёта и отлёта перелётных птиц, — наличие кормовой базы. Немалое значение для сроков миграций имеет также длина пути; в отдельных случаях птицы прилетают поздно потому, что зимовали в отдалённых местах .
Причины перелётов птиц заключаются в сезонных изменениях окружающей среды. Эти изменения приводят к тому, что птицы совершают миграции длиной в тысячи километров к местам, где экологическая обстановка в это время больше соответствует их требованиям. Благодаря сезонным миграциям птицы заселяют и используют для размножения такие места, которые в течение определённого периода года совершенно не пригодны для их существования .
Не все птицы совершают миграции при помощи перелётов. Большинство видов пингвинов регулярно мигрируют вплавь , иногда на тысячи километров. Некоторые морские птицы (тупики, кайры, чернозобая гагара ) мигрируют на зимовье из-за полярного круга, преодолевая бо́льшую часть пути вплавь. Голубой тетерев осуществляет регулярные миграции на различные высоты преимущественно пешком. Во время засухи длинные миграции пешком осуществляют и эму .

Многие виды птиц даже вне сезона размножения тяготеют к объединению в стаи, преимущественно одновидовые . Внутри стай отмечается т. н. социальное облегчение: если одна птица начинает чиститься, питаться, купаться и т. д., находящиеся рядом птицы вскоре начинают делать то же самое. В стаях часто также существует социальная иерархия: каждая особь имеет свой собственный ранг или «общественное положение», определяемое полом, размерами, силой, окраской и т. п. [ страница не указана 1212 дней ]

Более 200 видов птиц занимаются муравлением . Они отдыхают на муравьиных гнёздах, где муравьи, забравшись к ним на крылья и перья, удаляют эктопаразитов. Другие птицы «купаются» в муравейниках или давят муравьёв и смазывают ими оперение , преследуя аналогичные цели .
Кочевые муравьи , например Eciton burchellii , привлекают птиц, следующих за муравьями , таких как древолазовые или полосатые муравьеловки , которые используют муравьёв как загонщиков насекомых и других мелких членистоногих . Наиболее известны таким поведением представители семейства полосатых муравьеловок ( Thamnophilidae ), хотя оно описано и у древолазовых, мухоловковых, кардиналовых, кукушковых, дроздовых, совиных, бюльбюлевых и в некоторых других семействах.
Некоторые птицы специализируются на похищении корма, добытого другими птицами ( клептопаразитизм ). Чаще всего это отмечается у морских птиц, основу рациона которых составляет рыба. Особенно склонны к клептопаразитизму фрегаты и поморники .
Среди птиц весьма широко распространён комменсализм . Целый ряд видов млекопитающих и птиц нередко объединяются на время кормёжки в смешанные группы, порою существующие продолжительное время. Широко известны связи птиц с травоядными млекопитающими, населяющими открытые пространства. Так, стада копытных в Африке сопровождают буйволовые скворцы . В подобных случаях птицы ловят насекомых, потревоженных млекопитающими, а также склёвывают кровососущих насекомых и кожных паразитов непосредственно с сопровождаемых зверей .
Медоуказчики , чтобы добраться до содержимого улья, привлекают к нему медоеда , который, разрывая гнездо, поедает мёд, а птицы склёвывают личинок и соты.
На нескольких небольших островах Новой Зеландии в проливе Кука гаттерии часто поселяются в норах совместно с гнездящимися буревестниками. При этом птицы и рептилии живут вместе, не причиняя друг другу вреда .

Почти все птицы при нападении на них активно защищаются. Но специальные приспособления в этом направлении развиты не столько против внешних врагов, сколько против особей того же вида. Большая часть этих приспособлений выработалась в связи с драками самцов или самок в брачный период. Например, шпоры — костные и роговые наросты на цевке у куриных , на сгибе крыла у ряда гусеобразных и ржанкообразных .
При защите большинство птиц пользуется теми орудиями, которые используются ими при добывании пищи. Во многих случаях эта защита сопровождается устрашающим поведением: гуси злобно шипят, вертишейка ворочает вытянутой шеей и головой, напоминая змею, буревестники и качурки изрыгают на нападающего дурно пахнущую маслянистую жидкость, некоторые хищные птицы забрызгивают нападающего своими экскрементами, выпь и филин , поднимая перья, увеличивают размеры тела и принимают угрожающую позу .

Более пассивные способы защиты сводятся главным образом к покровительственной окраске, широко распространённой как у яиц, так и птенцов и взрослых птиц. Иногда нормальная защитная поза имитирует какой-нибудь предмет, например, высиживающий яйца серый лесной козодой сидит почти вертикально с вытянутой шеей, становясь чрезвычайно похожим на основание обломанной ветви. Не менее оригинальную защитную позу занимает исполинский белоног из Австралии .
Отдельные виды птиц используют химическую защиту от хищников. Некоторые трубконосые способны выделять из желудка неприятную маслянистую жидкость против агрессора . Обитающие в Новой Гвинее дроздовые мухоловки выделяют на кожу и перья сильнодействующий нейротоксин .
Некоторые птицы, гнездящиеся на земле, стараются увести хищников от гнезда при помощи отвлекающих манёвров. Кулики и некоторые воробьиные притворяются ранеными, имитируя ранение в крыло, волочат его за собой, бегают вокруг врага, подпуская его на минимальное расстояние. Так птица будет убегать до тех пор, пока не уведёт хищника на безопасное расстояние. После этого птица внезапно взлетает и возвращается к оставленному гнезду .

Большинство видов птиц так или иначе страдает от паразитов и инфекционных заболеваний . Наиболее распространённые у птиц паразитические организмы представлены разнообразными группами клещей , пухоедов и вшей (например, Ricinidae и пухопероеды ), гельминтов, а также микропаразитами, грибками, простейшими ( птичья малярия ), бактериями и вирусами ( псевдочума птиц , птичий грипп ). Среди вызываемых клещами птичьих болезней дерманиссиоз , кнемидокоптоз , ламинозиоптоз , стерностомоз и цитодитоз . По крайней мере 2500 видов клещей из 40 семейств ( Pterolichidae , Pteronyssidae , Xolalgidae и другие) живут в тесной взаимосвязи с птицами, занимают характерные для них места на перьях, коже и даже на клюве . Эти клещи могут иметь только форетические отношения, но нарушают покой хозяев и провоцируют недоедание . Среди истинных паразитов такие клещи, как Dermanyssus и Ornithonyssus ( Dermanyssoidea ) . Три надсемейства астигматных клещей ( Analgoidea , Pterolichoidea и часть Pyroglyphoidea ) образуют парафилетическую группу специализированных перьевых клещей (около 2000 видов, более 440 родов, 33 семейства), связанных с птицами . Эндемик Антарктиды блоха Glaciopsyllus antarcticus паразитирует на антарктическом буревестнике . Куриный клещ переносит ряд заболеваний (в том числе энцефалит кур), снижает яйценоскость кур, приводит их к истощению и анемии . Летучая мышь белокрылый вампир питается кровью птиц .
Среди микроорганизмов известны возбудители бактериальных, вирусных, грибковых и протозойных заболеваний у птиц . Наиболее опасными принято считать инфекционные заболевания, которые протекают у птиц очень быстро, а некоторые из них представляют большую опасность и для людей ( птичий грипп , H5N1 , орнитоз ). Среди микозов птиц наиболее часто встречаются кандидамикоз , аспергиллёз , кожные инфекции, вызываемые грибками родов и .

Птицы являются важными составляющими разнообразных экосистем , входят в пищевые цепи и оказывают непосредственное воздействие на экосистемы . Особенно заметна роль птиц в экосистемах островов. Поскольку птицы заселяли острова намного быстрее млекопитающих, они там способны полностью выполнять экологическую роль млекопитающих .
Хищные и насекомоядные птицы — естественные регуляторы численности организмов, которыми они питаются. В свою очередь, многие виды птиц служат пищей для целого ряда позвоночных животных .
Некоторые птицы, питающиеся нектаром, являются важными опылителями растений ( орнитофилия ) . Орнитофилия характерна для более чем ста семейств цветковых растений; в ней участвует около 2000 видов птиц из 50 семейств — среди них особенно заметную роль играют колибри , цветочницы , нектарницы , медососы , попугаи лори и др . В отдельных случаях птицы являются единственными опылителями растений. Поэтому у растений и птиц-опылителей нередко наблюдается коэволюция .
Многие виды, питающиеся плодами, играют важную роль в распространении семян ( орнитохория ). Семена многих видов растений прорастают только после того, как прошли через пищеварительный тракт птиц .
Ещё на ранних этапах развития человечества люди обладали знаниями о птицах, их образе жизни, местах массовых гнездовий и имели для птиц названия. Об этом свидетельствуют фольклорные источники, летописи и архивные документы. Однако последовательное и планомерное изучение птиц началось только в XVIII веке с началом множественных экспедиций .
На территории стран СССР первый этап в изучении птиц связан с именем Петера Симона Палласа и его экспедиций в 1768—1775 гг. Во второй половине XIX века Михаил Мензбир организовал новое интенсивное изучение птиц России, в ходе которого были собраны значительные коллекции птиц. В советский период исследования птиц ещё более расширились. Большое значение имела научно-педагогическая деятельность Михаила Мензбира, Петра Сушкина , Сергея Бутурлина , Льва Бёме , Владимира Флинта и многих других орнитологов-фаунистов. Одной из главных задач советских орнитологов была работа над составлением видовых и подвидовых списков фауны СССР . В этот период выпускается несколько многотомных публикаций, посвящённых птицам Советского Союза.
Современная орнитология — одна из наиболее развитых отраслей зоологии, тесно связанная с рядом биологических дисциплин .
Отношения между человеком и птицами имеют давнюю историю и разнообразную природу. В большинстве случаев человек использует птиц для получения мяса, перьев и пуха, а также в религиозных обрядах, для развлечений и в целях коммуникации ( почтовые голуби ). Иногда между человеком и птицами наблюдаются мутуалистические взаимоотношения, например кооперация в поиске мёда между медоуказчиками и африканскими народами, например, борана . В других случаях отношения между человеком и птицами можно рассматривать как комменсализм , например, со стороны домашнего воробья , который зависит от человеческой деятельности. Некоторые виды птиц могут наносить значительный ущерб сельскому хозяйству и представляют угрозу для самолётов . В то же время деятельность человека (охота, загрязнение окружающей среды, разрушение биотопов, хищничество со стороны домашних кошек, собак и т. д.) приводит к гибели множества птиц .
Наблюдение за полётом птиц подтолкнуло людей к изобретению первых летательных аппаратов, а дальнейшее его изучение продолжает оказывать влияние на развитие современной авиации .
Птицы могут участвовать в переносе на большие расстояния многих заболеваний. Известно более 40 инфекционных и инвазионных заболеваний, общих для человека и птиц, либо переносимых последними ( птичий грипп , сальмонеллёз , орнитоз , кампилобактериоз , лямблиоз и др.). От птиц к человеку могут передаваться вирусы гриппа, например H5N1 , хотя эта передача низкоэффективна . Большую роль в поддержании природных очагов заболеваний играют болотные и околоводные птицы, а также хищные птицы, питающиеся мышевидными грызунами . Многие виды диких птиц являются хозяевами личинок, нимф и взрослых иксодовых и гамазовых клещей , а также блох, передающих человеку и другим млекопитающим ряд вирусных и бактериальных трансмиссивных заболеваний . Птицы могут также переносить гельминтов и ряд протозойных заболеваний (кокцидиозы, опирохетозы).
В психиатрии описана специфическая фобия у людей — , заключающаяся в навязчивом страхе птиц .


Человеком одомашнено несколько видов птиц с целью получения от них яиц , мяса , пера и пуха , а также для использования в религиозных обрядах , для развлечения и в целях коммуникации ( почтовый голубь ) .
С давних времён различные птицы играли важную роль в питании человека, и сегодня остаются одним из основных источников животного белка. Разведение домашних птиц составляет отдельную отрасль сельского хозяйства — птицеводство . Для получения мяса птицы разводят мясные породы кур, уток, гусей, домашних индеек, цесарок, а также перепелов, фазанов, страусов и мясных голубей. Наряду с домашними птицами человек употребляет в пищу многие дикие виды птиц, не относящиеся к пернатой дичи , добываемой на охоте. Например, древние греки ели лесных голубей, галок, сов и чаек, ввозили из Африки фламинго. Во Франции XVI века голубей употребляли в пищу наряду с другими дикими птицами (например, кроншнепами и белыми цаплями). В эти же времена синицы, чибисы, дрозды, малиновки, зяблики, воробьи, жаворонки и сойки считались отличной едой по всей Европе. Сегодня певчие птицы составляют заметную часть рациона жителей Азии, Африки, Латинской Америки и отдельных районов Средиземноморья, при этом практически не употребляются в пищу в других регионах. Например, в Китае и Иране популярными остаются жареные воробьи; а коренные племена Амазонии употребляют в пищу попугаев .
Побочной продукцией птицеводства являются пух и перо (используются в качестве изоляции в одежде и постельных принадлежностях), а отходы производства используются для изготовления мясо-костной муки . Особый тип промыслового использования птиц — добыча гуано (помёта морских птиц), которое используется в качестве ценного органического удобрения . Места его добычи ограничены исключительно засушливыми областями тропиков и приурочены к островкам западного берега Южной Америки .
Куры — наиболее распространённый среди птиц объект лабораторных исследований ; они используются для исследований морфогенеза , анализа мутационных процессов , составления карт сцепления генов и т. п. В биологии развития в качестве модельного организма широко используется и немой перепел . На зебровой амадине исследуют генетику поведения, механизмы обучения и прочее . Куриный эмбрион — классическая культуральная среда в вирусологии .
Многие виды одомашнены с практической целью и в качестве домашних животных — певчих и декоративных птиц. Наиболее популярными являются попугаи: волнистые попугайчики , кореллы, какаду , жако , ара и другие. Канарейки благодаря яркой окраске и мелодичному пению стали одними из самых популярных певчих птиц, содержащихся в неволе. Распространено содержание ряда других воробьинообразных певчих птиц — различных видов амадин , щеглов, чижов и т. п. Относительно популярными во всём мире остаются голуби . На садовых участках и парках в качестве декоративных птиц могут содержаться павлины , куры , различные виды фазанов, а на прудах — белые и чёрные лебеди , различные виды уток ( мандаринка , огарь и др.) .
В 13 районах Японии , а также в ряде мест Китая , местным населением практикуется ловля рыбы японскими бакланами .
Почтовых голубей использовали многие культуры Средиземноморья задолго до нашей эры . Затем они активно использовались в Древнем Риме , Персии , Китае и Индии . Позднее голубиная почта стала весьма популярной во всём мире, и до изобретения телеграфа в середине XIX века была самым быстрым способом передачи сообщений. Она оставалась важным способом коммуникации вплоть до окончания Второй мировой войны .
В малозаселённых районах США для выявления утечек газа на газопроводах в природный газ добавляют вещества с запахом тухлого мяса и затем наблюдают за кружащими над трубопроводами грифами-индейками . Увидев скопления птиц, обходчики обследуют эти места и выявляют повреждение труб .
Длительное время (в Великобритании вплоть до 1987 года ) канареек использовали для обнаружения в шахтах рудничного газа . Как животные с быстрым метаболизмом и интенсивным дыханием, они очень чувствительны к ядовитым или вытесняющим воздух газам, включая метан и угарный газ , и гибнут даже от незначительной примеси последнего в воздухе. Шахтёры брали клетку с канарейкой в шахту и во время работы следили за птицей. Канареек часто использовали и горноспасатели, спускавшиеся в аварийные шахты .
Местное население тихоокеанского архипелага Санта-Крус , принадлежащего Соломоновым Островам , вплоть до XX века использовало тевау — товарные деньги , представляющие собой 10-метровую полосу из растительных волокон, покрытую красными перьями поверх жёстких коричневых перьев голубя .
Различные виды ткачиковых в Африке и Центральной Азии в периоды своего массового размножения в отдельные годы могут уничтожать значительную часть урожая зерновых на полях. В Северной Африке и Испании скворцы существенно вредят урожаю винограда и косточковых культур. В Европе и Америке от ряда птиц страдают посевы различных огородных культур. Однако многие из видов птиц, которые могут наносить в определённые сезоны ущерб сельскому хозяйству, в другие сезоны и в других условиях истребляют насекомых-вредителей. Отдельные виды (например, голуби) своим помётом наносят вред памятникам и архитектурным сооружениям .

Основная угроза для птиц со стороны человека — разрушение среды обитания , хотя количественно оценить его последствия трудно . Главные антропогенные причины гибели птиц, для которых такие оценки существуют, — столкновения с окнами и стеклянными панелями зданий (по данным для США, от этого гибнут от 100 до 1000 млн птиц в год, что составляет от 0,5 до 5 % всех местных птиц) , с автомобилями (60—80 млн), коммуникационными вышками (4—50 млн), охота (120 млн), домашние кошки (118 млн), линии электропередач (0,1—174 млн) и пестициды (72 млн) .
Описано множество случаев локальной массовой гибели птиц при использовании человеком различных ядов, в том числе для борьбы с насекомыми-вредителями, грызунами и сорняками, случаи гибели водных птиц при загрязнении нефтью и другими веществами в местах сброса отходов различных производств и т. п. Луговым птицам большой ущерб наносит разорение гнёзд и гибель птенцов при сенокосе и других сельскохозяйственных работах. В окрестностях крупных городов из-за частого вспугивания птиц нарушается нормальный ход насиживания и выкармливания птенцов . В конце XIX — начале XX века модно было украшать дамские шляпы птичьими перьями и даже маленькими чучелами птиц, в связи с чем массовый промысел птиц ради перьев привёл некоторые виды (ряд цапель, многие райские птицы, ряд куропаток) на грань вымирания .
С 1600 года по настоящее время в результате деятельности человека (непосредственного истребления, уничтожения мест обитания, завоза хищников и т. п.) вымерло более 200 видов и подвидов птиц . Были и виды, исчезнувшие по естественным причинам: их количество оценивают в четверть всех вымерших за этот период видов птиц . Активная колонизация новых земель сопровождалась полным либо почти полным истреблением некоторых птиц. Например, в Северной и Центральной Америке европейские переселенцы истребили 31 вид птиц. Самые тяжёлые потери понесли островные фауны: на Маскаренских островах вымерло 86 % обитающих там видов птиц, на Гваделупе — 39 %, на островах Лэсон [ уточнить ] и Мидуэй — 60 %, на Гавайских островах — 60 % видов :433 . К примеру, в конце XVII века был истреблён маврикийский дронт , в середине XIX века — бескрылая гагарка , в начале XX века — странствующий голубь . По состоянию на 2020 год 1486 видов птиц имеют статус уязвимых , вымирающих или находящихся на грани исчезновения , а 1017 видов близки к уязвимому положению .

|

|

|

|
Существованию видов птиц могут угрожать как естественные факторы (природные катаклизмы, глобальные изменения климата), так и антропогенные (хищнический промысел, загрязнение окружающей среды, разрушение биотопов). Охрана птиц имеет научные, культурные и хозяйственные аспекты, охватывая широкий круг мероприятий, направленных на сохранение и увеличение популяций видов и мест обитания птиц в различных регионах Земли .
Первые общества по охране птиц были созданы ещё в XIX веке: в 1886 году в США организовано Национальное Одюбоновское общество , в 1893 году — Куперовское орнитологическое общество ; в 1889 году в Великобритании — Королевское общество защиты птиц . В 1922 году была создана старейшая международная организация по защите птиц и среды их обитания — Международный совет охраны птиц ( International Council for Bird Preservation ), переименованный в 1993 году в BirdLife International .
В 1868 году на съезде немецких сельских и лесных хозяев было впервые выдвинуто предложение о заключении двусторонних договоров между разными странами об охране «полезных» видов птиц. В 1902 году была составлена и в 1905 году вступила в силу Парижская конвенция по охране птиц, полезных в сельском хозяйстве. В 1950 году была составлена новая «Международная конвенция по охране птиц», которая вступила в силу только в 1963 году. Она впервые закрепила принципы защиты от истребления любых дико живущих видов птиц. Созданные позднее Конвенция о водно-болотных угодьях, имеющих международное значение в качестве места обитания водоплавающих птиц (1971), Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой уничтожения (1973), Боннская конвенция (1979) и различные группы региональных договоров и охранных постановлений существенно расширили международно-правовое регулирование данной сферы .
С 1894 года в США отмечается День птиц — праздник, постепенно ставший международным, цель которого — привлечь внимание властей и граждан страны к проблемам сохранения исчезающих видов птиц .
В России с 1993 года действует Союз охраны птиц . Существуют также Итальянская лига защиты птиц (1969) и Всепольское общество охраны птиц (1991) , Украинское общество охраны птиц (1994) , Белорусское общество «Охрана птиц Отечества» (1998) . Благодаря действию этих и других общественных и научных организаций и обществ ( МСОП , Всемирный фонд дикой природы , Мензбировское орнитологическое общество ) были собраны данные о численности и выработаны меры по охране редких и исчезающих видов птиц.
В Красную книгу СССР (1978) было включено 63 вида птиц, в Красную книгу России (2001) включено 126 видов птиц , в Красную книгу Украины (2009) включено 87 видов .


С древних времён человек охотился на различных животных, включая птиц. Одни делали это ради добычи пропитания, другие — перьев, третьи — ради развлечения, четвёртые — собирали коллекции . В наши дни охота представляет собой главным образом хобби и развлечение . Лишь в некоторых регионах, преимущественно среди коренного населения, она служит для добывания пищи . Виды птиц, собирательно называемые в охотничьей практике « пернатая дичь », являются наиболее популярным объектом любительской спортивной охоты . «Охотничьей», как правило, считается птица со вкусным мясом, хотя причисление того или иного биологического вида к дичи носит условный характер и зачастую вызвано традицией — так, во Франции и Италии дрозды считаются ценной дичью, а в России к дичи вообще не относятся .
Во многих странах охота на птиц регламентирована на законодательном уровне, введены особые квоты на отстрел, а охотникам выдаются специальные разрешения и лицензии . Как правило, любительская охота на птицу проводится с помощью гладкоствольного ружья . Стрельба ведётся дробью . Лишь иногда, при больших расстояниях и при охоте на очень крупную птицу (глухарь, дрофа, гуси), охотники применяют нарезное оружие малого калибра, но это разрешено далеко не во всех странах. При охоте часто используют охотничьих собак — легавых , спаниелей , лаек и т. д.
Существует охота на дичь, включая пернатую, с использованием специально обученных ловчих птиц. Такая охота традиционно именуется соколиной , хотя, кроме собственно соколов , могут использоваться и другие хищные птицы — кречеты , беркуты и др. Охота с ловчими птицами зародилась на Ближнем Востоке и в Центральной Азии 4000 лет назад и сохранилась до настоящего времени . Орлов учили охотиться на лисиц, волков и даже антилоп . В России соколиная охота была особенно популярной среди знати в XVII веке . В Средние века соколиная охота была также широко распространена в Западной Европе. В качестве дорогостоящего развлечения сейчас она популярна в некоторых странах Ближнего Востока . Охота с ловчими беркутами в настоящее время практикуется в горных районах Казахстана и Монголии .
В Египте ежегодно убивают 5,7 млн птиц. Здесь в массе гибнут домовые воробьи, лысухи, жуланы и иволги. В Италии — 5,6 млн птиц в год. Особенно велики потери зябликов, луговых коньков, и певчих дроздов. В Сирии — 3,9 млн в год. Здесь велики потери серых и черноголовых славок, и полевых жаворонков. В топ-10 попали также Греция, Франция, Хорватия и Албания. Наибольшую проблему представляют лов натяжными сетями и ловушками, приманивание птиц с помощью аудиозаписей. Страшно жесток метод лова с помощью клея, которым намазывают ветки деревьев, в результате чего прилипшие птицы в страданиях гибнут от жажды и перегрева .

|

|
|||||||||
|
Гор и Тот
|
||||||||||
С давних времён птицы играют различные, в том числе и знаковые роли в фольклоре , мифологии, религии и массовой культуре многих народов мира. Обычно птицы выступали обязательным элементом религиозно-мифологических систем и ритуалов, обладающим различными функциями. Птицы могли быть божествами; демиургами ; героями, превращёнными людьми; ездовыми животными для богов и героев; тотемными предками и т. п.
В религиях птицы обычно были посланниками богов, например, в культе Маке-маке острова Пасхи ; в скандинавской мифологии два ворона Хугин и Мунин шептали новости в уши бога Одина . Птицы как воплощение божества играли важную роль в мифах о сотворении мира : во многих из них фигурирует великая птица- демиург или птица — помощник божественного творца. Божества-соколы почитались под разными именами во многих областях Древнего Египта и Нубии , но все они, как правило, были связаны с небом и солнцем. Например божество-сокол — Гор ; бог Тот изображался в образе человека с головой ибиса . В образе птицы древними египтянами изображалась и одна из составляющих души — « ба », покидающая тело после смерти .

В шумеро-аккадской мифологии птичьи черты есть у Иштар , Ашшура , Адада и других божеств. Согласно верованиям шумеров, в птичьи перья были одеты и души умерших в подземном царстве богов. Птицы были среди атрибутов многих древнегреческих богов : у Афины — сова, у Зевса и Диониса — орёл, у Геры — павлин, аист и кукушка. В связи с этим и по другим причинам в честь птиц названы 9 из 88 современных созвездий .
Почитание священных птиц было широко распространено в Древнем Китае : журавль, павлин, петух относились к существам ян , символизируя долголетие и удачную судьбу. Ацтеки вели происхождение бога Уицилопочтли от колибри, изображающегося в образе человека с зелёным оперением на голове, а Кетцалькоатль сочетал в себе черты змея и птицы .
Птицы также выступали в качестве религиозных символов. Голуби фигурируют в христианстве (голубь Ноя , голубь как символ Святого Духа ). Павлин у дравидов Индии символизировал Мать-Землю .
У многих народов птицы олицетворяли душу человека. Это обнаруживается в древних культурах Египта, Двуречья , Китая, Греции, а также в Сибири ( эвенки , якуты ) и Южной Америке (индейцы бороро ). Переход души после смерти в тело птиц фигурирует во множестве легенд, в том числе морских, согласно которым души утонувших моряков кружат возле кораблей в образе чаек и других морских птиц.
Предсказания, осуществляемые по поведению и пению птиц, были широко распространены в Вавилоне и через этрусков достигли Древнего Рима , где играли весьма значительную роль, а жрецы- авгуры отличались высоким статусом в обществе. В Древней Греции подобные предсказания не получили распространения и считались смехотворными .






Образы птиц часто использовались в сказках (« Сказка о золотом петушке », « Гуси-лебеди » и др.), баснях («Журавль и цапля», «Лиса и журавль») и детских рассказах (« Серая Шейка », « Оранжевое горлышко », « Чудесное путешествие Нильса с дикими гусями » и др.), где они выступают главными либо второстепенными персонажами. Значительное место среди этих образов занимают фантастические пернатые. Например, в средневековом арабском фольклоре фигурировала птица Рух , способная уносить в когтях и пожирать слонов. В мифологиях разных культур существует Феникс — птица, способная сжигать себя и возрождаться из пепла . Широко распространён мотив превращения героев в птиц. Например, сказочные девы обращались в лебедей, уток или голубей, а ведьмы, колдуны и злые духи — в воронов, сов и т. п.


Образы птицы — одни из наиболее распространённых образов живых существ, которые встречаются в искусстве.
В качестве символа женской красоты, изящества и нежности лебедь предстал в балете Петра Чайковского « Лебединое озеро ». Выдающееся место среди ритуальных танцев занимают т. н. «птичьи» танцы: «утиный» танец, танцы орла и индюка у американских индейцев, «журавлиные» танцы в Греции, славянский вариант весеннего хоровода — « », литовская детская игра «уточка» и многие другие .
Художественные изображения птиц с давних времён существуют во многих мировых культурах. Ранние изображения птиц с сакральным смыслом датируются верхним палеолитом ( наскальная живопись и предметы «мобильного» искусства). К ним относятся и изображения с древнеегипетских фресок возрастом 3500 лет. В живописи итальянского Возрождения ( Рафаэль и др.) распространён образ мадонны со щеглом — символом страстей. Художники, посвятившие свои работы птицам, сыграли важную роль в увеличении интереса к пернатым. В XVII—XVIII вв. были изданы иллюстрированные труды по естествознанию Джона Рэя , Фрэнсиса Виллоби , Томаса Бьюика . В XIX веке Александр Уилсон , Джон Гульд и Джон Одюбон достигли выдающих успехов в изображении птиц. Работы Гульда и Одюбона считаются настоящими произведениями искусства, а не просто иллюстрациями . На их картинах птицы сочетаются с элементами их природного окружения. В России одним из первых зоологов-художников стал Николай Северцов , а Василий Ватагин считается основателем российского анималистического жанра . В китайской живописи с X века существовало направление под названием « цветы и птицы » , позаимствованное затем японцами и корейцами.
Изображения птиц, в том числе скульптурные, всегда были популярной темой в искусстве фарфора, росписи и вышивке тканей.

Птицы стали центральными образами ряда кинофильмов. Наиболее известен классический кинофильм Альфреда Хичкока « Птицы », в котором пернатые нападали на прохожих и врезались в окна и стены домов .
Различные, преимущественно антропоморфные, птицы часто становятся персонажами мультфильмов : « Возвращение блудного попугая », « Рио », « Камешек и пингвин », « Приключения пингвинёнка Лоло », « Вэлиант: Пернатый спецназ », « Пингвины Мадагаскара » и многие другие.
Птицы присутствуют на 35 % современных государственных гербов . Примерно половина из них включают изображения одноглавых и двуглавых орлов, а также сокола, ястреба, кондора; на семи гербах изображён голубь. Из архетипических мифических образов присутствуют грифоны , птица Хумай , Гаруда ( гербы Таиланда , Индонезии и Улан-Батора ), кречет Манаса , из национальных и традиционных символов — венценосный журавль , птица-секретарь , дронт , петух, кетцаль , морские птицы (страны Океании), ворон и др. Изображения птиц могут быть доминирующей эмблемой герба либо составной частью основополагающих эмблем .







Птицы часто встречаются в современной культуре. Их изображения в том или ином виде нередки на обложках научных книг о природе и художественных произведений; на рекламных плакатах и обложках музыкальных дисков, в музыкальных видеоклипах, на керамических и ювелирных изделиях, предметах быта. Изображения птиц часто помещают на почтовых марках и монетах . Метафора « белая ворона » обозначает лицо, имеющее поведение или систему ценностей, отличные от других лиц общества.
В США целый ряд птиц был избран символами отдельных штатов .
В странах Юго-Восточной Азии пользуются популярностью « петушиные бои » — состязания боевых петухов , которых стравливают друг с другом, ставшее своеобразным азартным видом спорта, в котором зрители делают ставки. В Великобритании популярными остаются голубиные гонки , в ходе которых специально обученные стаи голубей, соревнуясь между собой, должны пролететь большие расстояния .
|
|
|
|||
|---|---|---|---|---|
| Словари и энциклопедии |
|
|||
| Таксономия |
|
|||
|
|
||||
|
|
Эта статья победила на конкурсе
статьи года
и была признана статьёй 2015 года русской Википедии.
|




























