Interested Article - Гистондеацетилаза 4

- 2021-03-04
- 2
Локус
:
![]()
Гистондеацетила́за 4 ( англ. Histone deacetylase 4, HDAC4 ) ( КФ ) — белок , кодируемый у человека геном HDAC4 , расположенным на 2-й хромосоме . Как и все ферменты группы гистондеацетилаз , близкой к сиртуинам , гистондеацетилаза 4 катализирует удаление с остатков лизина в N-концевой части коровых гистонов ( , , H3 и ), что изменяет структуру хроматина . Де гистонов является одним из механизмов транскрипционной и эпигенетической регуляции, оказывает влияние на ход клеточного цикла и участвует в регуляции развития . Работа HDAC4 регулируется путём различных посттрансляционных модификаций и взаимодействий с разнообразными белками, иногда тканеспецифичными. Нарушение работы HDAC4 приводит к развитию многих заболеваний, в том числе раковых , поэтому ингибиторы HDAC4 могут иметь важное медицинское применение.
Ген и регуляция экспрессии
У человека ген HDAC4 расположен на 2-й хромосоме (2q37.3) , имеет длину около 353,49 килобаз (кб), содержит 37 экзонов и даёт начало 8980 мРНК -транскриптам. У мыши гомологичный ген Hdac4 имеет длину около 215,7 кб, располагается на 1-й хромосоме и даёт начало 3960 мРНК-транскриптам. HDAC4 экспрессируется в различных тканях , и уровень экспрессии зависит от интенсивности различных стимулов. Несмотря на огромное количество процессов, регулируемых HDAC4, и уникальные механизмы регуляции активности этого белка, о механизмах регуляции его экспрессии известно мало. Транскрипционные факторы Sp1 и Sp3 связываются непосредственно со специфическими GC-обогащёнными участками в промоторе HDAC4 и запускают транскрипцию HDAC4 . HDAC4 не экспрессируется в ядрах эмбриональных стволовых клеток мыши, однако при начале дифференцировки клеток уровень его экспрессии резко возрастает .
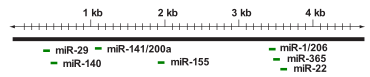
Показано, что в регуляции экспрессии HDAC4 участвуют несколько микроРНК , среди которых miR-1, miR-29, miR-140, miR-155, miR-200a, miR-206 и miR-365, действующие в клетках разных типов. miR-200a непосредственно связывается с 3'-нетранслируемой областью (3'-UTR) мРНК HDAC4 и подавляет его экспрессию. miR-1 специфична для клеток мышц и стимулирует , действуя на 3'-UTR мРНК HDAC4 и подавляя экспрессию HDAC4 . Белок mTOR контролирует MyoD-зависимую транскрипцию miR-1 через вышележащий энхансер , а опосредуемая miR-1 репрессия HDAC4 приводит к образованию и последующему слиянию миоцитов . Временная трансфекция прогениторных клеток кардиомиоцитов при помощи miR-1 и miR-499 снижала скорость пролиферации и вызывала усиленную дифференцировку человеческих прогениторных клеток кардиомиоцитов и эмбриональных стволовых клеток в кардиомиоциты через репрессию HDAC4 . Кроме того, miR-22, подавленная при гепатоцеллюлярной карциноме , подавляет пролиферацию и склонность к образованию опухолей через регуляцию HDAC4 .
Кроме того, сверхэкспрессия miR-206 и miR-29 подавляла экспрессию HDAC4 на уровне трансляции как в присутствии, так и в отсутствие трансформирующего фактора роста-бета (TGF-β) через взаимодействие с 3'-UTR HDAC4 . Экспрессия miR-206 и miR-29, задействованных в дифференцировке клеток мышц, отрицательно регулируется TGF-β, поэтому обработка миогенных клеток TGF-β вызывает повышенную экспрессию HDAC4. miR-29b выступает ключевым регулятором дифференцировки в остеобласты , действуя на белки HDAC4, TGF-β3, ACVR2A, CTNNBIP1 и DUSP2. miR-140, специфичная для хрящевой ткани , непосредственно действует на 3'-UTR HDAC4 . Мыши, лишённые miR-140, имеют карликовый фенотип из-за нарушения развития хондроцитов . miR-365, активируемая при механическом воздействии, связана с модуляцией дифференцировки хондроцитов, действуя непосредственно на HDAC4 . У , имеющих человеческую miR-155, miR-155 действует на HDAC4 и нарушает регуляцию транскрипции гена 6 в В-клетках . Искусственно усиленная экспрессия HDAC4 в человеческих клетках В-клеточной лимфомы уменьшала индуцируемую miR-155 пролиферацию и усиливала апоптоз . Всё это свитедельствует о важной роли микроРНК, специфично действующих на HDAC4 , в модуляции клеточного ответа и биологических функций клеток разных типов в ответ на различные стимулы .
Структура
Структура доменов

Человеческий ген HDAC4 кодирует белки длиной от 972 до 1084 аминокислотных остатков, а мышиный гомолог Hdac4 — от 965 до 1076 аминокислотых остатков. HDAC4 содержит уникальный регуляторный домен на N-конце , который взаимодействует с различными транскрипционными факторами, и цинк -содержащий каталитический домен на С-конце . Анализ кристаллической структуры показывает, что правильно фолдированный (уложенный) цинк-связывающий домен необходим для формирования репрессорного комплекса. N-концевой участок мономера HDAC4 консервативен и содержит обогащённый глутамином домен (19 остатков глутамина из 68), который укладывается в прямую альфа-спираль , участвующую в сборке тетрамера гистондеацетилазы 4. Тетрамер HDAC4 не имеет регулярно расположенных неполярных аминокислотных остатков и протяжённого . Вместо этого взаимодействие между субъединицами обеспечивается множеством гидрофобных островков, расположенных внутри участков с полярными аминокислотными остатками, а глутамин-обогащённые участки принимают участие в сворачивании альфа-спиралей мономеров и их взаимодействии друг с другом . С-концевой цинк-связывающий домен играет ключевую роль в распознавании субстрата и связывании HDAC4 с репрессорным комплексом HDAC3-NCoR. Детальный анализ кристаллической структуры показал, что между цистеином 669, расположенным в цинк-связывающем домене, и цистеином 700 соседней молекулы может формироваться межмолекулярная дисульфидная связь .
Посттрансляционные модификации
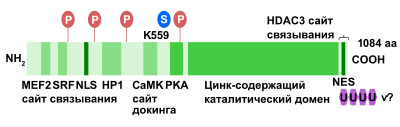
Посттрансляционные модификации HDAC4 могут изменять его внутриклеточную локализацию и состав взаимодействующих с ним белков. Хорошо известно, что одной из ключевых функций HDAC4 является репрессия транскрипции генов-мишеней через регуляцию конденсации и структуры хроматина. Недавние исследования показали критическую роль посттрасляционных модификаций в контроле клеточных ответов, в которых задействована HDAC4. Было показано, что HDAC4 может фосфорилироваться, сумоилироваться, карбонилироваться, убиквитинироваться и разрезаться под действием различных ферментов .
Фосфорилирование
Фосфорилирование /дефосфорилирование обеспечивает быструю и эффективную репрессию гистондеацетилаз (HDAC) класса IIа, к которому принадлежит и HDAC4. Обратимое фосфорилирование — регуляторный механизм, необходимый для функционирования HDAC4. HDAC4 взаимодействует с семейством белков 14-3-3 , которые специфично связываются с фосфо серин -содержащими консервативными мотивами . Фосфорилирование этих остатков серина создаёт сайты связывания с шапероном семейства 14-3-3, который сопровождает фосфорилированную HDAC4 при транспорте из ядра в цитоплазму . HDAC4 может фосфорилироваться следующими белками: , ERK1/2 , протеинкиназа А (РКА) и .
Стимуляция CaMK запускает миогенез путём разрушения комплексов -HDAC и последующего экспорта HDAC из ядра. специфично связывается с HDAC4 через уникальный сайт докинга . Фосфорилирование HDAC4 по остаткам серина S246, S467 и S632 при помощи CaMKII усиливает ядерный экспорт и предотвращает ядерный импорт HDAC4 с последующей репрессией генов-мишеней HDAC4. Передача сигнала через эндогенный CaMKII необходимо для индуцированного агонистом накопления HDAC4 в цитозоле кардиомиоцитов. Однако РКА фосфорилирует HDAC4 и регулирует протеолиз HDAC4 по остатку тирозина 207, а также выступает антагонистом CaMKII-опосредованной активации MEF2 путём регулирования протеолиза HDAC4. Продукт разрезания HDAC4, включающий в себя N-конец бывшего белка, селективно ингибирует активность MEF2, но не (SRF), выступая антагонистом CaMKII, но не влияя на выживание кардиомиоцита. Активация сигнального пути Ras - MAPK при экспрессии онкогенного белка Ras или в случае конститутивно активной MAPK/ERK-киназы 1 вызывает накопление HDAC4 в ядре миобластов. GSK3 может фосфорилировать HDAC4 по позициям 298 и 302, что приводит к разрушению HDAC4 протеасомами ; таким образом, этот белок выступает в качестве важного регулятора стабильности HDAC4 .
Аналогичным образом, важную роль в регуляции HDAC4 играют дефосфорилирующие ферменты — протеинфосфатазы . В условиях in vitro HDAC4 дефосфорилируется PP2A , которая сначала взаимодействует с N-концом HDAC4, а потом дефосфорилирует её. Регулируя дефосфорилирование HDAC4 по нескольким остаткам серина, в том числе и тем, которые входят в сайт связывания с белками 14-3-3, а также остатку серина 298, PP2A контролирует ядерный импорт HDAC4 .
Карбонилирование
Карбонилирование , или алкилирование, — характерная посттрансляционная модификация в клетках, подвергшихся окислительному стрессу . Карбонилированием называется ковалентное присоединение активной карбонильной группы к тиольной группе остатков цистеина в белке-субстрате. В ответ на стимулы, индуцирующие образование активных форм кислорода в клетке, в белке DnaJb5 остатки цистеина 274 и 276, а в HDAC4 — 667 и 669 окисляются и формируют внутримолекулярные дисульфидные связи, которые затем могут быть восстановлены тиоредоксином -1. Восстановление остатков цистеина 274 и 276 белка DnaJb5 необходимо для взаимодействия DnaJb5 и HDAC4, а восстановление остатков цистеина 667 и 669 HDAC4 подавляет её ядерный экспорт независимо от степени фосфорилированности .
Сумоилирование
Сумоилирование — это ковалентное присоединение к остаткам лизина белка. Как и при убиквитинировании , присоединение белков SUMO ( , и ) к остаткам лизина в белках-субстратах играет критическую роль в модуляции активности и разрушения этих белков. Было показано, что HDAC4 распознается SUMO1 по единственному остатку лизина (лизин-559), по которому и происходит сумоилирование. Оно осуществляется E3 SUMO-протеинлигазой и не влияет на внутриклеточное распределение HDAC4, а также на его взаимодействие с некоторыми из белков, с которыми она обычно взаимодействует. Однако HDAC4 с мутацией в позиции 559 значительно хуже функционирует и осуществляет репрессию транскрипции генов-мишеней по сравнению с диким типом . Сумоилированию HDAC4 препятствует её фосфорилирование CaMK4 .
Убиквитинирование
Обычно полиубиквитинирование направляет белки на деградацию протеасомами, в то время как моноубиквитинирование может иметь различные биологические эффекты. Убиквитинирование и протеасомное разрушение HDAC4 регулируется путём фосфорилирования , однако механизм и биологическое значение убиквитинирования HDAC4 ещё не выяснены .
Протеолиз
Перемещение HDAC4 между ядром и цитоплазмой также находится под влиянием протеолиза, которое происходит в процессе апоптоза. HDAC4 разрезается и по остатку аспартата 289. N-концевой фрагмент HDAC4, отрезаемый каспазами, содержит сигнал ядерной локализации и накапливается в ядре, репрессируя транскрипцию и вызывая гибель клетки, а также выступая сильным репрессором MEF2C. По сравнению с другими ядерными формами HDAC4, ядерный фрагмент, отрезаемый каспазами, вызывает гибель клетки и оказывает мощное подавляющее действие на - или SRF-зависимую транскрипцию, несмотря на то что не содержит С-концевого цинк-связывающего домена, необходимого для распознавания субстрата и связывания с корепрессорным комплексом HDAC3 -N-CoR. Создаваемый каспазами фрагмент слабо связывается с хроматином , в то время HDAC4, мутантная по сайту связывания 14-3-3, формирует более стабильные комплексы с белком HDAC5 .
Действие на клеточном уровне
Гистоны играют важнейшую роль в регуляции экспрессии генов. Ацетилирование/деацетилирование гистонов изменяет структуру хроматина и влияет на доступ факторов транскрипции к ДНК . HDAC4 относится к классу II семейства гистондезацетилаз/acuc/apha. Он обладает активностью гистондеацетилазы и подавляет транскрипциию, связываясь с промотором. Этот белок не связывает ДНК напрямую, а лишь посредством транскрипционных факторов и . Как и для всех гистондеацетилаз, для работы HDAC4 необходимы ионы Zn 2+ .
Как было рассмотрено выше, экспрессия гена HDAC4 может регулироваться на транскрипционном и посттранскрипционном (при помощи микроРНК и регуляции стабильности мРНК) уровнях, а также на уровне стабильности белка (разрушение протеазами). HDAC4 перемещается между ядром и цитоплазмой, а также выступает в качестве ядерного корепрессора , регулирующего развитие костей и мышц. Активность HDAC4 регулируется при помощи двух основных механизмов: внутриклеточной локализации и образования многобелковых комплексов с другими белками .
Внутриклеточное распределение
Как обсуждалось выше, перемещение HDAC4 между ядром и цитоплазмой может регулироваться при помощи посттрансляционных модификаций. Транслокация HDAC4 также регулируется через взаимодействие с транспортным фактором экспортином 1 , также известным как CRM1 , который управляет экспортом из ядра клеточных белков, имеющих обогащённый лейцином (NES). Кроме того, (Nup155), главный компонент ядерного порового комплекса (NPC), задействован в перемещении белков между цитоплазмой и ядром. Считается, что HDAC4 функционирует как корепрессор транскрипции, деацетилируя нуклеосомные гистоны. Поскольку деацетилазы гистонов не взаимодействуют непосредственно с ДНК, в настоящее время считается, что их привлечение к специфическим промоторам обеспечивают , распознающие определённые последовательности нуклеотидов в ДНК. HDAC4 также взаимодействует с различными белками, например, , гистонметилтрансферазой , различными транскрипционными факторами, что определяет функции этого белка в различных тканях (список белков, с которыми взаимодействует HDAC4, ). Имеется множество доказательств того, что гистондеацетилазы, и HDAC4 в их числе, деацетилируют не только гистоны, но и другие белки, в том числе различные транскрипционные факторы, что может служить регуляторным механизмом биологических сигнальных путей. Цитоплазматические функции HDAC4 хорошо изучены и освещаются ниже .
Регуляция деацетилирования гистонов
HDAC4 осуществляет деацетилирование как гистоновых, так и путём удаления с субстратов ацетильных групп цинк-содержащим каталитическим доменом. Обратимое ацетилирование по N-концевым лизиновым остаткам гистона 3 (позиции 9, 14, 18 и 23) и гистона 4 (позиции 5, 8, 12 и 16) вызывает деконденсацию нуклеосом, изменяет взаимодействие гистонов с ДНК и увеличивает доступность ДНК для транскрипционных факторов. Состояние ацетилированности гистонов управляется двумя группами белков противоположного действия: гистонацетилтрансферазами (НАТ), ацетилирующими гистонами, и гистондеацетилазами, осуществляющими их деацетилирование. В отличие от HDAC6, HDAC4 и HDAC5 взаимодействует с HDAC3 и RbAp48. Каталитический домен HDAC склонен к образованию многобелкового комплекса с корепрессорным комплексом SMRT-NCoR-HDAC3. Целостность каталитического домена HDAC4 необходима для привлечения корепрессорного комплекса HDAC3-N-CoR и его дальнейшей деацетилазной активности. Как деацетилаза HDAC4 не активна в отсутствие связывания с HDAC3 .
Регуляция деацетилирования негистоновых белков
Белок Runx2 служит главной мишенью сигнального пути BMP . Сигнальный путь BMP-2 стимулирует ацетилирование Runx2, опосредованное . Эта модификация увеличивает активность Runx2 и ингибирует -опосредованную деградацию Runx2. HDAC4 и HDAC5 деацетилируют Runx2, позволяя этому белку подвергнуться Smurf-опосредованному разрушению. Ингибирование HDAC увеличивает ацетилирование Runx2, усиливает дифференцировку остеобластов, стимулированную сигнальным путём , и увеличивает образование костей. Недавние исследования показали, что HDAC4 может деацетилировать такие цитоплазматические белки, как , MEKK2 и STAT1 .
Деметилирование гистонов
Ацетилирование и метилирование гистонов — наиболее полно изученные эпигенетические метки. Триметилирование по позициям H3K4, H3K36 или H3K79 приводит к тому, что хроматин принимает активную форму, характерную для эухроматина . Эухроматин также характеризуется высокой степенью ацетилирования гистонов. Поэтому HDAC могут убирать эпигенетические метки, репрессируя транскрипцию. Метилированный H3K9 создаёт сайт связывания для хромодомен -содержащего белка HP1, который индуцирует репрессию транскрипции и переход эухроматина в гетерохроматин . HDAC4 участвует в эпигенетической регуляции генов через взаимодействие с H3K9- SUV39H1 и HP1, обеспечивая эффективный механизм сайленсинга генов-мишеней MEF2 путём осуществления и деацетилирования, и метилирования. Деметилирование H3K9 тесно связано с перемещением HDAC4 между цитоплазмой и ядром. Особенно значительно триметилирование H3K9 в условиях стресса в промоторе 5'- ацетилхолинэстеразы (AChE), и накопление такой гистоновой метки ассоциируют с рекрутированием SUV39H1 и HP1 на промотор (AChE) .
Кроме того, HDAC4 отрицательно регулирует транскрипционный фактор MEF2 через взаимодействие с SUMO Е2-конъюгирующим ферментом Ubc9. Сверхэкспрессия HDAC4 приводила к чрезмерному сумоилированию MEF2 в условиях in vivo . HDAC4 стимулирует сумоилирование MEF2 по тому же остатку лизина, который ацетилирует коактиватор MEF2, ацетилтрансфераза CREBBP , так что, возможно, ацетилирование и сумоилирование MEF2 взаимодействуют при регуляции его активности. Впрочем, эта модель является предметом споров, и необходимы дополнительные эксперименты, чтобы выяснить, сумоилирует ли HDAC4 MEF2 непосредственно или же привлекает для этого SUMO E2-конъюгирующий фермент .
Физиологические функции
HDAC4 выполняет важнейшие функции в регуляции транскрипции генов, росте клеток, пролиферации и выживании, поэтому нарушения в экспрессии или работе этого белка приводят к развитию рака .
Костная и хрящевая ткани
HDAC4, экспрессируемая в прегипертофических хондроцитах, регулирует гипертрофию хондроцитов и эндоклональное образование костей, взаимодействуя с и ингибируя активность Runx2 — транскрипционного фактора, необходимого для гипертрофии хондроцитов. У мышей , нокаутных по HDAC4, наблюдается преждевременное окостенение развивающихся костей из-за преждевременной эктопической гипертрофии хондроцитов; похожий фенотип проявляется у особей, в хондроцитах которых Runx2 экспрессируется постоянно. Runx2 может ацетилироваться белком р300, и ацетилированная форма Runx2 предотвращает убиквитинирование белков. HDAC4 и HDAC5 играют противоположные роли, деацетилируя Runx2 и позволяя белкам разрушаться по Smurf-зависимому пути. TGF-β подавляет дифференцировку остеобластов, действуя на HDAC4 и HDAC5, которые в дифференцирующихся остеобластах через взаимодействие со рекрутируются в комплекс Smad3/Runx2, располагающийся на последовательности ДНК, связывающейся с Runx2. Сверхэкспрессия HDAC4 стимулирует TGF-β1 -индуцированный в стволовых клетках синовиального слоя , однако подавляет гипертрофию в дифференцировавшихся из них хондроцитах .
Мышечная ткань
Первая стадия миогенеза включает формирование миобластов, которые экспрессируют особый набор транскрипционных факторов, в том числе MEF2C. У мышей, лишённых MEF2C, наблюдаются нарушения в морфогенезе сердца , и на стадии образования петли в развитии сердца развитие организма прекращается. HDAC4 связывается непосредственно с MEF2, ингибируя его функционирование, и регулирует дифференцировку клеток мезодермы в кардиомиобласты, подавляя экспрессию GATA4 и . Обработка ингибиторами HDAC вызывает спецификацию клеток мезодермы в будущие кардиомиоциты, о чём можно судить по повышению содержания в них транскриптов Nkx2-5, MEF2C, GATA4 и сердечного α- актина . Таким образом, HDAC подавляют дифференцировку мезодермальных клеток в кардиомиоциты. Сверхэкспрессия HDAC4 подавляет кардиомиогенез, о чём свидетельствует снижение уровня экспрессии генов, отвечающих за развитие кардиомиоцитов .
Показано, что при дифференцировке мышечных клеток HDAC4 управляет репрессией генов, рекрутируя MEF2 к промоторам репрессируемых генов. Репрессия транскрипции генов MEF-2/HDAC-комплексом обусловлено CaMK-индуцированной транслокацией HDAC4 и HDAC5 в цитоплазму. В сердцах трансгенных мышей со сверхэкспрессией активного наблюдалась гипертрофия сердца с повышением содержания некоторых эмбриональных транскриптов, например, предсердного натрийуретического фактора , и значительное повышение активности MEF2C .
Все сокращения скелетной мускулатуры контролируются нервной системой . HDAC4 в норме накапливается в области . Утрата иннервации вызывает сопутствующее накопление HDAC4 в ядре мышечной клетки и уменьшение экспрессии генов, регулируемых MEF2. При хирургической денервации или в случае нервномышечного заболевания бокового амиотрофического склероза повышенный уровень HDAC4 необходим для эффективной репрессии MEF2-зависимых структурных генов. Усиленная экспрессия HDAC4 оказывает эффект, схожий с эффектом денервации, и активирует транскрипцию эктопического ацетилхолинового рецептора ( nAChR ) по всему мышечному волокну. Инактивация HDAC4 предотвращает индуцированную денервацией транскрипцию синаптических рецепторов nAChR и . Особенно много HDAC4 содержится в ядрах быстрых окислительных скелетных мышечных волокнах, и нокаут HDAC4 усиливает гликолиз в мышечных трубочках .
Нервная система
HDAC4 имеется в околоядерной области цитоплазмы большинства нейронов , однако его локализация в ядре варьирует. В зубчатой извилине ядерной экспрессии HDAC4 не наблюдается, в то время как в ядрах нейронов из других зон содержат HDAC4. В норме HDAC4 локализуется в цитоплазме нейронов головного мозга и выращенных в культуре гранулярных нейронов мозжечка . HDAC4 быстро переносится в ядро в ответ на низкий уровень калия и опасный уровень глутамата , которые индуцируют смерть нейрона. Обработка фактором выживания нейронов BDNF препятствует ядерной локализации HDAC4, в то время как ингибитор CaMK, стимулирующий апоптоз, способствует накоплению HDAC4 в ядре. Более того, локализованного в ядре HDAC4 стимулирует апоптоз нейронов и подавляет функционирование в качестве факторов транскрипции белков MEF2 и CREB. Гистондеацетилазы играют важную роль в выживании нейронов и развитии фоторецепторов . Транскрипционный комплекс MEF2-HDAC4 участвует в выживании нейронов и является мишенью . Внутриклеточная локализация HDAC определяется активностью нейрона. Спонтанная электрическая активность необходима для ядерного экспорта HDAC4, но не HDAC5 .
Поджелудочная железа
Показано, что HDAC4, HDAC5 и HDAC9 (класс IIa HDAC) проявляют неожиданно ограниченную экспрессию эндокринных β- и δ-клеток поджелудочной железы . Эти HDAC выступают ключевыми регуляторами β/δ-клеток поджелудочной железы. Анализ мышей, мутантных по HDAC класса IIa, показал, что количество инсулин -продуцирующих β-клеток повышено у мышей, нокаутных по HDAC5 и HDAC9, а у нокаутных по HDAC4 и HDAC5 — соматостатин -продуцирующих δ-клеток. Сверхэкспрессия HDAC4 и HDAC5 приводила к снижению количества β- и δ-клеток .
Клиническое значение
Сердечно-сосудистые заболевания
Гипертрофия сердца — это ответ сердца на различные внешние и внутренние стимулы, которые приводят к биомеханическому стрессу. Множество сердечно-сосудистых заболеваний , в том числе инфаркт миокарда , артериальная гипертензия и различные изменения в сократимости сердца, обусловлены мутациями саркомерных белков, причём эти мутации заставляют взрослое сердце увеличиваться в размерах из-за гипертрофического роста кардиомиоцитов. В кардиомиоцитах CaMKII-зависимое фосфорилирование HDAC4 приводит к гипертрофическому росту, которое может блокироваться при HDAC4, не воспринимающей какие-либо сигналы. Исследования мышей, лишённых miR-22, показали, что miR-22 необходима для гипертрофического разрастания сердца в ответ на стресс и непосредственными мишенями этой микроРНК являются HDAC4 и Sirt1 .
Кроме того, HDAC4 участвует в регуляции сокращения миофиламентов через регуляцию деацетилирования MLP. HDAC4, HAT и p300/CREBBP-ассоциированный фактор ( PCAF ) связаны с сердечными миофиламентами. HDAC4 и PCAF связаны с Z-дисками и I- и A-полосами сердечных саркомеров. MLP, белок, ассоциированный с Z-дисками, функционирует как сенсор механического натяжения сердца, и в ацетилированном виде является мишенью HDAC4 и PCAF .
Неврологические заболевания
Болезнь Хантингтона (HD) — это нейродегенеративное генетическое заболевание, при котором нарушается координация мышечных сокращений, возникают когнитивные расстройства и психиатрические проблемы. Показано, что в случае HD miR-22 может иметь многосторонний антинейродегенеративный эффект, включающий в себя ингибирование апоптоза и влияние на гены (в том числе HDAC4, RCOR1 и ), задействованных в развитии HD .
Недостаточная экспрессия HDAC4 при развитии сетчатки приводит к апоптозу палочек и биполярных интернейронов (BP), в то время как сверхэкспрессия снижает по сравнению с нормой количество погибающих BP-клеток. Кроме того, у мышей с дегенерирующей сетчаткой сверхэкспрессия HDAC4 удлиняло срок жизни фоторецепторов. Эффект выживания был обусловлен активностью HDAC4 в цитоплазме .
Дефекты HDAC4 могут служить причиной синдрома брахидактилии с отсталостью умственного развития. Физические проявления этого синдрома напоминают таковые наследственной остеодистрофии Олбрайта . Среди этих симптомов — умеренные лицевые нарушения, врождённые пороки сердца , брахидактилия типа Е, умственная отсталость, отставание в развитии, , расстройства аутистического спектра , коренастое телосложение. Изучение 278 пациентов с шизофренией и контрольных здоровых 234 людей из корейской популяции анализ однонуклеотидных полиморфизмов показал, что ген HDAC4 связан с развитием шизофрении. — это нейродегенеративное заболевание, обусловленное мутацией в гене Atm . У мышей, дефектных по этому гену, накопление HDAC4 в ядре приводило к нейродегенерации .
Рак
В некоторых случаях острых лейкозов хромосомная транслокация , приводящая к слиянию гена PLZF , кодирующего белок PLZF , с геном, кодирующим рецептор ретиноевой кислоты , даёт начало химерному белку PLZF-RARα, который, как считают, осуществляет конститутивную репрессию генов, ответственных за дифференцировку. Было установлено, что HDAC4 взаимодействует с лейкозным белком PLZF-RARα и управляет репрессией генов дифференцировки в лейкозных клетках. Подавление активности HDAC ингибиторами HDAC в клинических и фундаментальных исследованиях показало потенциальную пользу HDAC в лечении рака. Белок BCL6 отвечает за выживание и/или дифференцировку при В-клеточной лимфоме, возникающей из-за хромосомных перестроек. HDAC4 связывается с BCL6 и PLZF in vivo и in vitro и через них управляет репрессией транскрипции. Показано, что микроРНК miR-155, сверхэкспрессия которой чаще всего наблюдается в опухолях и при злокачественных гематологических заболеваниях, может непосредственно связываться с 3'-UTR HDAC4 и подавлять её трансляцию. Эктопическая экспрессия HDAC4 в клетках В-клеточной лимфомы человека приводила к снижению miR-155-индуцированной пролиферации и усилению апоптоза .
Наибольшая экспрессия HDAC4 наблюдается в пролиферативной части нормального эпителия тонкой и толстой кишки , и при дифференцировке её экспрессия снижается. HDAC4 взаимодействует с Sp1 и убирает ацетильные группы с гистона Н3 в сайте связывания Sp1/Sp3 на проксимальном промоторе белка р21 , репрессируя транскрипцию. Индукция этого промотора путём сайленсинга HDAC4 останавливало рост раковых клеток и подавляло рост опухоли в модели человеческой глиобластомы . Х -связанный супрессор опухолей FOXP3 необходим для экспрессии р21 в нормальном эпителии, и недостаток FOXP3 приводит к отрицательной регуляции р21, что происходит в некоторых случаях рака молочной железы . FOXP3 специфично ингибируется при связывании HDAC4 и локальном увеличении ацетилирования гистона Н3. При гепатоцеллюлярной карциноме HDAC4 непосредственно регулируется miR-22. Более того, в ткани гепатоцеллюлярной карциномы при отрицательной регуляции miR-22 уровень HDAC4 повышался. Кроме того, в клетках этой опухоли HDAC4 является также мишенью miR-200a .
При раке яичника нередко наблюдается устойчивость к платиновой химиотерапии , причём показано, что в устойчивых опухолях наблюдается повышенная экспрессия HDAC4. PLU-1/ , экспрессия которого повышается в некоторых случаях рака молочной железы , взаимодействует с HDAC4 и экспрессируется совместно с ним в раковых клетках этого типа. Показано, что в образцах ткани здорового мочевого пузыря для HDAC4-положительных образцов была значительно ниже, чем в образцах опухоли мочевого пузыря . Кроме того, содержание HDAC4 при переходных карциномах мочевого пузыря значительно выше, чем в нормальных тканях. HIF1α — необходимая часть транскрипционного комплекса HIF-1, который регулирует ангиогенез , клеточный метаболизм и может отвечать за развитие рака. Ацетилирование HIF1α положительно регулируется shРНК HDAC4 , но не shРНК HDAC1 или HDAC3. Ингибирование HDAC4 снижает и транскрипционную активность HIF-1, и экспрессию ряда генов-мишеней HIF-1, а также уменьшает устойчивость к химиотерапии с применением доцетаксела . Установлено, что HDAC4 может быть задействована в развитии остеосаркомы и рака толстой кишки . , препарат, предназначенный для лечения устойчивой к удалению опухоли предстательной железы , непосредственно связывается с HDAC4, тем самым ингибируя деацетилирование гистонов и транскрипционные факторы, зависящие от HDAC4, например, HIF-1α .
Ингибиторы
К настоящему моменту известно множество , относящихся к различным группам соединений. Среди них есть гидроксаматы ( трихостатин А , вориностат ), ( , ), алифатические кислоты ( бутират , , вальпроевая кислота ), и его производные. Перечисленные ингибиторы неспецифичны и подавляют работу всех HDAC, а не только HDAC4. Их применение может оказаться перспективным при лечении различных раковых заболеваний . Известны и специфичные ингибиторы HDAC4, в частности, производные трифлуорометил-1,2,4-оксидазола. Эти соединения могут оказаться эффективными при лечении болезни Хантингтона, и диабета .
Взаимодействия с другими белками
HDAC4, как было выявлено, взаимодействует с:
| Белок | Комментарий | Источники |
|---|---|---|
| BCL6 | Может связываться не только с HDAC4, но и с другими родственными HDAC класса IIa: HDAC5 и HDAC7 | |
| BTG2 | Может также связываться с HDAC1 | |
| GATA1 | HDAC ингибируют этот белок. Также он взаимодействует с HDAC3 и HDAC5 | |
| HDAC3 | Вместе входят в состав репрессорного комплекса HDAC3-NCoR | |
| MAPK1 | Локализация HDAC4 зависит от сигнального пути Ras-MAPK | |
| MAPK3 | Локализация HDAC4 зависит от сигнального пути Ras-MAPK | |
| Ингибируется HDAC4 | ||
| Ингибируется HDAC4 | ||
| Вместе входят в состав репрессорного комплекса HDAC3-NCoR | ||
| NCOR2 | Вместе входят в состав репрессорного комплекса HDAC3-NCoR |
Примечания
- ↑ - Ensembl , May 2017
- ↑ - Ensembl , May 2017
- Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ↑ Grozinger C. M. , Hassig C. A. , Schreiber S. L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1999. — Vol. 96, no. 9 . — P. 4868—4873. — .
- Fischle W. , Emiliani S. , Hendzel M. J. , Nagase T. , Nomura N. , Voelter W. , Verdin E. (англ.) // The Journal of biological chemistry. — 1999. — Vol. 274, no. 17 . — P. 11713—11720. — .
- ↑ . Дата обращения: 6 июня 2015. 11 января 2018 года.
- ↑ Wang Z. , Qin G. , Zhao T. C. (англ.) // Epigenomics. — 2014. — Vol. 6, no. 1 . — P. 139—150. — doi : . — .
- . Дата обращения: 3 октября 2017. 9 июня 2019 года.
- Guo L. , Han A. , Bates D. L. , Cao J. , Chen L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2007. — Vol. 104, no. 11 . — P. 4297—4302. — doi : . — .
- .
- Dokmanovic M. , Clarke C. , Marks P. A. (англ.) // Molecular cancer research : MCR. — 2007. — Vol. 5, no. 10 . — P. 981—989. — doi : . — .
- Abdel-Magid A. F. (англ.) // ACS medicinal chemistry letters. — 2013. — Vol. 4, no. 8 . — P. 692—693. — doi : . — .
- Lemercier C. , Brocard M. P. , Puvion-Dutilleul F. , Kao H. Y. , Albagli O. , Khochbin S. (англ.) // The Journal of biological chemistry. — 2002. — Vol. 277, no. 24 . — P. 22045—22052. — doi : . — .
- Farioli-Vecchioli S. , Tanori M. , Micheli L. , Mancuso M. , Leonardi L. , Saran A. , Ciotti M. T. , Ferretti E. , Gulino A. , Pazzaglia S. , Tirone F. (англ.) // FASEB journal : official publication of the Federation of American Societies for Experimental Biology. — 2007. — Vol. 21, no. 9 . — P. 2215—2225. — doi : . — .
- Watamoto K. , Towatari M. , Ozawa Y. , Miyata Y. , Okamoto M. , Abe A. , Naoe T. , Saito H. (англ.) // Oncogene. — 2003. — Vol. 22, no. 57 . — P. 9176—9184. — doi : . — .
- ↑ Fischle W. , Dequiedt F. , Hendzel M. J. , Guenther M. G. , Lazar M. A. , Voelter W. , Verdin E. (англ.) // Molecular cell. — 2002. — Vol. 9, no. 1 . — P. 45—57. — .
- Grozinger C. M. , Schreiber S. L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2000. — Vol. 97, no. 14 . — P. 7835—7840. — doi : . — .
- Fischle W. , Dequiedt F. , Fillion M. , Hendzel M. J. , Voelter W. , Verdin E. (англ.) // The Journal of biological chemistry. — 2001. — Vol. 276, no. 38 . — P. 35826—35835. — doi : . — .
- ↑ Zhou X. , Richon V. M. , Wang A. H. , Yang X. J. , Rifkind R. A. , Marks P. A. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2000. — Vol. 97, no. 26 . — P. 14329—14333. — doi : . — .
- ↑ Miska E. A. , Karlsson C. , Langley E. , Nielsen S. J. , Pines J. , Kouzarides T. (англ.) // The EMBO journal. — 1999. — Vol. 18, no. 18 . — P. 5099—5107. — doi : . — .
- Lemercier C. , Verdel A. , Galloo B. , Curtet S. , Brocard M. P. , Khochbin S. (англ.) // The Journal of biological chemistry. — 2000. — Vol. 275, no. 20 . — P. 15594—15599. — doi : . — .
- ↑ Huang E. Y. , Zhang J. , Miska E. A. , Guenther M. G. , Kouzarides T. , Lazar M. A. (англ.) // Genes & development. — 2000. — Vol. 14, no. 1 . — P. 45—54. — .
Литература
- Verdin E. , Dequiedt F. , Kasler H. G. (англ.) // Trends in genetics : TIG. — 2003. — Vol. 19, no. 5 . — P. 286—293. — doi : . — .
- Grozinger C. M. , Hassig C. A. , Schreiber S. L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1999. — Vol. 96, no. 9 . — P. 4868—4873. — .
- Miska E. A. , Karlsson C. , Langley E. , Nielsen S. J. , Pines J. , Kouzarides T. (англ.) // The EMBO journal. — 1999. — Vol. 18, no. 18 . — P. 5099—5107. — doi : . — .
- Wang A. H. , Bertos N. R. , Vezmar M. , Pelletier N. , Crosato M. , Heng H. H. , Th'ng J. , Han J. , Yang X. J. (англ.) // Molecular and cellular biology. — 1999. — Vol. 19, no. 11 . — P. 7816—7827. — .
- Youn H. D. , Grozinger C. M. , Liu J. O. (англ.) // The Journal of biological chemistry. — 2000. — Vol. 275, no. 29 . — P. 22563—22567. — doi : . — .
- Grozinger C. M. , Schreiber S. L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2000. — Vol. 97, no. 14 . — P. 7835—7840. — doi : . — .
- Wang A. H. , Kruhlak M. J. , Wu J. , Bertos N. R. , Vezmar M. , Posner B. I. , Bazett-Jones D. P. , Yang X. J. (англ.) // Molecular and cellular biology. — 2000. — Vol. 20, no. 18 . — P. 6904—6912. — .
- Zhang C. L. , McKinsey T. A. , Lu J. R. , Olson E. N. (англ.) // The Journal of biological chemistry. — 2001. — Vol. 276, no. 1 . — P. 35—39. — doi : . — .
- McKinsey T. A. , Zhang C. L. , Lu J. , Olson E. N. (англ.) // Nature. — 2000. — Vol. 408, no. 6808 . — P. 106—111. — doi : . — .
- Zhou X. , Richon V. M. , Wang A. H. , Yang X. J. , Rifkind R. A. , Marks P. A. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2000. — Vol. 97, no. 26 . — P. 14329—14333. — doi : . — .
- McKinsey T. A. , Zhang C. L. , Olson E. N. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2000. — Vol. 97, no. 26 . — P. 14400—14405. — doi : . — .
Ссылки
- 2021-03-04
- 2