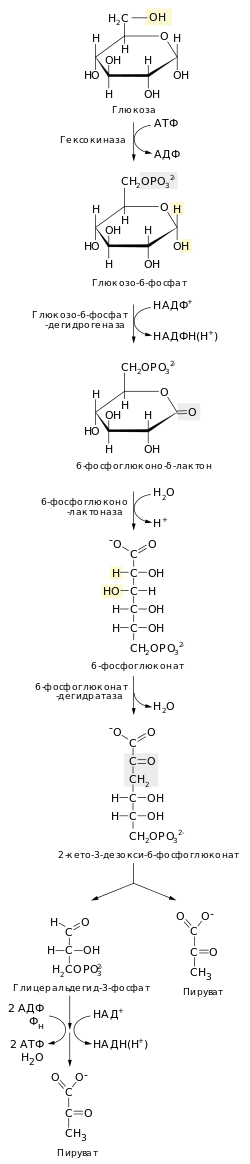Военно-морские силы Индии
- 1 year ago
- 0
- 0

Локус
:
![]()
Нуклеофозми́н ( англ. Nucleophosmin , также известен как нуматри́н , NPM1 , NPM , NO38 , я́дрышковый фосфопротеи́н B23 ) — ядрышковый белок , у человека кодируется геном NPM1 , локализованным на 5-й хромосоме . Нуклеофозмин перемещается между ядром и цитоплазмой и действует как многофункциональный шаперон нуклеиновых кислот , принимающий участие в таких процессах, как биогенез рибосом , ремоделирование хроматина , регуляция митоза , поддержание стабильности генома , репарация ДНК и транскрипция . Нарушения в работе нуклеофозмина могут приводить к развитию злокачественных новообразований и других заболеваний; в частности, мутации , затрагивающие его ген, приводят к развитию острого миелоидного лейкоза .
Нуклеофозмин был впервые описан вместе с нуклеолином (С23) Busch и коллегами в 1973 году .
У человека ген NPM1 располагается на 5-й хромосоме в локусе 5q35.1 и содержит 12 экзонов . Известно более дюжины псевдогенов этого гена . Человеческий нуклеофозмин (основная изоформа ) состоит из одной полипептидной цепи длиной 294 аминокислотных остатка и имеет массу 32 575 дальтон . Нуклеофозмин высококонсервативен среди таких организмов , как человек, грызуны , куры и рыбы . Известны три изоформы нуклеофозмина: полноразмерная NPM1.1, NPM1.2, образующаяся в результате альтернативного сплайсинга , и NPM1.3. мРНК NPM1.2 и NPM1.1 отличаются по 3'-концевому экзону, причём NPM1.2 короче основной изоформы (состоит из 259 аминокислотных остатков). О функциях и особенностях экспрессии недавно описанной изоформы NPM1.3 известно мало, однако установлено, что у неё отсутствует внутренний участок в С-концевом домене .
Нуклеофозмин относится к семейству белковых шаперонов гистонов типа нуклеоплазмина (NPM-белков), к которому, кроме него, принадлежат ещё два белка (NPM2 и ). Эти белки обнаружены у млекопитающих , рыб, птиц , мух , но не у бактерий и дрожжей . Отличительной чертой NPM-белков является наличие консервативного N-концевого домена олигомеризации . Эти белки функционируют как и могут собираться в декамеры при расположении двух пентамеров друг над другом с образованием структуры, напоминающей сэндвич. Считается, что в таком виде они способны связываться с гистонами . Показано, что для сборки пентамеров дисульфидные связи , образуемые остатками цистеина , не играют существенной роли . В 2014 году было также показано, что N-концевой домен олигомеризации проявляет структурный полиморфизм при переходе между разными конформационными состояниями NPM1: от высокоупорядоченного пентамера до неупорядоченного мономера . Равновесие между мономерной и пентамерной формами NPM1 регулируется его фосфорилированием и связыванием с другими белками . Так, фосфорилирование смещает равновесие в сторону мономеров .
NPM1 содержит участки, необходимые для олигомеризации, шаперонной активности, связывания с нуклеиновыми кислотами, а также ядерной локализации. Функционированию NPM1 как шаперона вполне соответствует чрезвычайная термическая и химическая стабильность его N-концевого домена . Первые 15 аминокислот составляют участок, обогащённый метионином , однако функциональное значение этого участка неизвестно. Возможно, он нужен для усиления инициации трансляции , поскольку метиониновые кодоны входят в хорошие последовательности Козак . Хотя этот богатый метионином участок не является необходимой частью белка, он может оказывать влияние на его конформацию. NPM1 содержит три участка, обогащённых отрицательно заряженными (кислыми) аминокислотными остатками (А1, А2 и А3). Возможно, кислые участки играют роль в нейтрализации заряда белка. Кор NPM1, содержащий А1, может в одиночку слабо связываться с гистонами и , однако для связывания с гистонами и необходимы кислые участки А2 и А3 (их отрицательный заряд имитирует отрицательный заряд сахарофосфатного остова ДНК и РНК ). С-концевой домен NPM1 содержит группы положительно заряженных (основных) аминокислот, за которыми следует участок, обогащённый аминокислотами с ароматическими радикалами. Этот участок участвует в связывании с нуклеиновыми кислотами, связывании с АТР , переносе гистонов, обладает рибонуклеазной активностью и содержит сигнал ядрышковой локализации (NoLS) . Кроме того, С-концевой домен нуклеофозмина способен специфично распознавать G-квадруплексы в ДНК .
В 2015 году было показано, что α-спираль Н2 (остатки 264—277) из С-концевого домена нуклеофозмина может формировать токсичные амилоидоподобные агрегаты с фибриллярной структурой при физиологических условиях .
На рисунке ниже представлена схема строения NPM1.
| Олигомеризация | Основный участок | ||||||||||
| Met | A1 | A2 | NLS | A3 | HeD/DBD | NoLS | |||||
| 1 − | − 294 | ||||||||||
Нуклеофозмин подвергается таким посттрансляционным модификациям , как фосфорилирование, , убиквитинирование и . NPM1 считается ядрышковым фосфопротеином и может фосфорилироваться несколькими киназами — такими, как (CKII), polo-подобная киназа 2 (Plk2), и комплекс / CDK2 . Фосфорилирование нуклеофозмина оказывает влияние на его активность, олигомеризацию, способность к перемещениям внутри ядра и клетки в целом, а также локализацию в клетке, в частности, фосфорилирование может усиливать его сродство к компонентам рибосомы и, таким образом, влиять на биогенез рибосом. Так, CKII фосфорилирует NPM1 по остатку S 125, который находится в одном из кислых участков. Этот участок необходим для работы NPM1 как шаперона, и фосфорилирование по S125 вызывает диссоциацию субстрата от NPM1 . Фосфорилирование по этому остатку, помимо того, уменьшает способность NPM1 к внутриклеточным и внутриядерным перемещениям. Фосфорилирование казеинкиназой II усиливает связывание NPM1 с сигналами ядерной локализации (NLS) и белка ВИЧ . Конкретные сайты сумоилирования NPM1 точно не установлены, но сумоилирование NPM1 может влиять на его локализацию и стабильность. может убирать «метку» SUMO с NPM1. Убиквитиновые «метки» с NPM1 удаляет фермент , стабилизируя NPM1, в то время как отсутствие USP36 приводит к дефектам в биогенезе рибосом. Интересно, что NPM1 может сам доставлять USP36 в ядрышко путём непосредственного связывания . Ацетилирование NPM1 посредством оказывает двойной эффект: оно стимулирует перемещение NPM1 из ядрышка в нуклеоплазму и, помимо этого, необходимо для стимуляции РНК-полимераза II -зависимой транскрипции посредством NPM1. При ВИЧ-инфекции , вызванной ВИЧ-1, уровень ацетилирования NPM1 повышается .
NPM1 курсирует между ядром и цитоплазмой и содержит сигналы как ядерного импорта (NLS), так и (NES). Перемещение NPM1 между ядром и цитоплазмой необходимо для осуществления некоторых его функций, в частности, для экспорта из ядра и контроля удвоения центросом . NPM1 может доставлять в ядрышко небольшие основные белки. В частности, он может связываться с вирусными и ядрышковыми белками Rev, Rex, и и способствовать их локализации в ядрышке. В ядре NPM1 располагается в основном в ядрышке, хотя некоторое его количество присутствует в нуклеоплазме. В ходе митоза он обнаруживается в остатках ядрышка в перихромосомном слое и в области митотического веретена . В ядрышке NPM1 находится в основном в гранулярном компоненте, где созревают прерибосомные частицы, а также на границе плотного фибриллярного компонента .
Последовательности и молекулярные механизмы, обеспечивающие ядрышковую локализацию NPM1, не вполне ясны, однако известен ряд ключевых моментов. Так, мутации, разрушающие структуру мономеров и олигомеров, заметно снижают накопление этого белка в ядрышке. Кроме того, показано, что для ядрышковой локализации NPM1 совершенно необходимы два остатка триптофана W288 и W290, которые, предположительно, обеспечивают правильную вторичную структуру для связывания с нуклеиновыми кислотами и тем самым облегчают связывание. Показано, что для ядрышковой локализации и стабильности NPM1 необходимы также два остатка лизина K263 и K267. NMP1 определённо содержит сигнал ядерной локализации, однако имеются разногласия относительно того, какой именно мотив в центральном участке выполняет эту роль. Изоформа NPM1.2 обнаруживается в клетках в небольших количествах, причём в цитоплазме и нуклеоплазме, что свидетельствует в пользу необходимости С-конца для ядрышковой локализации . В настоящее время считается, что механической основой удержания нуклеофозмина в ядрышке является сильное связывание его С-конца с G-квадруплексами в области рДНК .
Рассматриваются несколько возможных кандидатов на роль сигнала ядерного экспорта, который у NPM1 располагается в домене олигомеризации. Первый из них — последовательность 42- L S L R T V SL-49, где мутации в положениях L42A и L44A блокируют ядерный экспорт NPM1. Второй мотив — 94- I T P PVVLRL-102, где мутация L102A блокирует не только ядерный экспорт, но и вообще перемещение NPM1 между ядром и цитоплазмой .
Большое влияние на внутриклеточную локализацию NPM1 оказывает малая GTPаза Rac1 . В клетках, экспрессирующих активную Rac1, NPM1 перемещается из ядра в цитоплазму. Впрочем, NPM1 способен отрицательно регулировать Rac1 .
Нуклеофозмин имеет множество разнообразных клеточных функций, которые подробно освещены ниже.
NPM1 обладает признаком белковых шаперонов: он связывается с денатурированными белковыми субстратами. В условиях in vitro он препятствует агрегации и тепловой денатурации некоторых белков. NPM1 может связываться с прерибосомными частицами (в частности, ) и поэтому может выполнять роль фактора сборки рибосом. В условиях in vitro он способствует разрезанию пре- рРНК и функционирует как эндорибонуклеаза , обеспечивающая созревание транскрипта рРНК. NPM1, кроме того, участвует в осуществлении контроля качества созревающих рРНК . Нокдаун NPM1 при помощи малых интерферирующих РНК нарушает процессинг пре-РНК (в частности, в 28S рРНК ), а блокирование его перемещения между ядром и цитоплазмой подавляет экспорт рибосомных субъединиц, что приводит к снижению скорости роста клеток. NPM1 может непосредственно взаимодействовать с рядом рибосомных белков, в частности, RPL5, и . NPM1 образует комплекс с другим белком своего семейства, NPM3, причём NPM3 негативно регулирует активность NPM1 при биогенезе рибосом. Интересно, что варианты NPM1, лишённые домена связывания с нуклеиновыми кислотами, также подавляют биогенез рибосом подобно NPM3. Таким образом, NPM1 способствует росту и пролиферации клеток, участвуя в нескольких стадиях биогенеза рибосом .
В условиях in vitro NPM1 может собирать нуклеосомы и деконденсировать ДНК сперматозоидов . Есть подтверждения функционирования NPM1 в ядрышке в качестве шаперона гистонов. Любопытно, что in vitro NPM3 подавляет способность NPM1 к сборке нуклеосом .
Ядрышковый супрессор опухолей (далее ARF) является одним из важнейших белков, с которыми связывается NPM1. Повышение количества ARF в клетке препятствует перемещению NPM1 между ядром и цитоплазмой, способствует его деградации и замедляет созревание 28S рРНК. В нормальных условиях NPM1 способствует ядрышковой локализации и стабильности ARF .
NPM1 задействован в процессах репликации , транскрипции, рекомбинации и репарации ДНК. Он может участвовать в ремоделировании хроматина, воздействуя на сборку нуклеосом или регулируя модификации гистонов посредством привлечения соответствующих ферментов .
NPM1 связывается с белком ретинобластомы (pRB) и в условиях in vitro стимулирует работу ДНК-полимеразы α , поэтому он может влиять на репликацию ДНК. Кроме того, in vitro NPM1 стимулирует репликацию ДНК аденовируса .
NPM1 непосредственно участвует в регуляции транскрипции ДНК на нескольких уровнях. Во-первых, он связывается с промоторами , с которыми взаимодействует белок c- Myc , и стимулирует транскрипцию, опосредованную РНК-полимеразой II. NPM1 участвует в регуляции кругооборота c-Myc и потому может влиять на рост и злокачественное перерождение клеток. Во-вторых, NPM1 взаимодействует с — негативным регулятором РНК-полимеразы II и облегчает транскрипцию. В-третьих, NPM1 при условии ацетилирования коровых гистонов увеличивает темпы транскрипции. Ацетилирование NPM1 (с образованием ацетилированной формы Ac-NPM1) приводит к разрушению нуклеосом и активации транскрипции. Ac-NPM1 встречается главным образом в нуклеоплазме в связанном с РНК-полимеразой II виде. Однако в ходе митоза NPM1 связывается с GCN5 и подавляет GCN5-опосредованное ацетилирование свободных и мононуклеосомных гистонов. В-четвёртых, NPM1 может выступать как корепрессор или коактиватор транскрипции, связываясь с , IRF1 , p53 , NF-κB и другими транскрипционными факторами . Например, было показано, что NPM1 участвует в транскрипционном ответе на ретиноевую кислоту в миелоидных клетках . В ходе дифференцировки, запущенной ретиноевой кислотой, NPM1 образует комплекс с активирующим транскрипционным фактором 2α и функционирует как корепрессор, привлекая деацетилазы гистонов . В-пятых, NPM1 участвует в регуляции транскрипции генов в ядрышке, и активирует транскрипционный фактор TAF(I)48, контролирующий транскрипцию генов рРНК. Активность РНК-полимеразы I жёстко регулируется несколькими супрессорами опухолей (p53) и онкогенами (c-Myc). Поскольку и c-Myc, и NPM1 связываются с ядрышковым хроматином в области рДНК и могут активировать транскрипцию, опосредованную РНК-полимеразой I, вполне может быть, что сверхэкспрессия NPM1 усиливает синтез рРНК, запущенный c-Myc (подобно тому, как эти два белка активируют транскрипцию с промоторов, с которыми работает РНК-полимераза II). Это явление имеет важное значение в контексте регуляции клеточного роста и злокачественной трансформации. Для связывания NPM1 с ядрышковым хроматином необходимы РНК-связывающая способность NPM1 и ядрышковый транскрипционный фактор UBTF . Более того, NPM1 способствует ядрышковой локализации терминирующего фактора РНК-полимеразы I . Таким образом, NPM1 играет важную роль в транскрипции, опосредованной РНК-полимеразами I и II .
Показано, что фосфорилированный NPM1 привлекается к участкам ДНК, повреждённым излучением. Подавление транскрипции рДНК и процессинга рРНК в отсутствие повреждений ДНК вызывает быстрое перемещение ядрышкового белка NPM1 в нуклеоплазму. Отмечалось, что уровни мРНК и белка NPM1 (а также его способность к связыванию РНК) значительно повышались при повреждениях ДНК, вызванных УФ-излучением . Усиленная экспрессия NPM1 делает клетки более устойчивыми к УФ-индуцированной гибели. По-видимому, NPM1 функционирует как гистоновый шаперон во время или после репарации двуцепочечных разрывов в ДНК . Кроме того, NPM1 регулирует стабильность, активность и накопление в ядрышке белков, осуществляющих эксцизионную репарацию оснований .
Сумоилирование — это посттрансляционная модификация, которая заключается в ковалентном присоединении небольших к другим белкам, что изменяет их работу в разнообразных клеточных процессах, в том числе апоптозе , , регуляции транскрипции, стабильности белков и репарации ДНК. «Метка» SUMO удаляется с белка под действием SUMO-деконъюгирующей протеазы (SENP). SENP3 и SENP5 локализуются в ядрышке и связываются с NPM1, поэтому NPM1 может принимать участие в регуляции сумоилирования. Нокдаун NPM1 и нокдаун ядрышковых белков SENP приводит к схожим дефектам в биогенезе рибосом .
У мышей , гетерозиготных по Npm1 , наблюдались некоторые нарушения митоза, а именно неограниченная репликация центросом и геномная нестабильность. Установлено, что некоторое количество белка NPM1 обнаруживается в районе веретена деления в ходе метафазы . В веретене NPM1 располагается вместе с белком . Находящийся в веретене NPM1 модифицирован (в частности, фосфорилирован). NPM1 может непосредственно участвовать в удвоении центросом в некоторых клетках. Это подтверждается тем, что NPM1 связывается с неудвоенными центросомами в ходе интерфазы и покидает их при фосфорилировании комплексом cdk2/циклин E по остатку T199, что запускает удвоение центросом. Впрочем, у мышей фосфорилирование NPM1 по остатку T198 происходит в ходе всего клеточного цикла . Фосфорилирование по T199 увеличивает сродство NPM1 к протеинкиназе , что, в свою очередь, увеличивает активность ROCK II. Нокдаун ROCK II препятствует удвоению центросом, а постоянная экспрессия активной формы фермента способствует ей. Кроме того, располагаясь рядом и между двумя центриолями одной неудвоенной центросомы, NPM1 соседствует с белком Crm1 , который принимает участие в регуляции удвоения центросом и сборки веретена деления. Подавление работы Crm1 приводит к увеличению содержания циклина Е в центросоме, диссоциации NPM1 и началу дупликации центросомы. Показано также, что NPM1 связывается с митотическими центросомами и, по-видимому, через взаимодействие с комплексом /Crm1 подавляет их повторное удвоение. После фосфорилирования cdk2 NPM1 покидает митотические центросомы .
Показано, что NPM1 связывается с центромерным белком , который замещает гистон Н3 в области центромер. Поэтому NPM1 может играть роль в поддержании стабильности центромер. В быстрорастущих клетках HeLa недостаток NPM1 приводил к остановке митоза из-за непрохождения веретена деления и активации p53. В этих клетках наблюдались нарушения формирования митотического веретена и удвоения центросом .
NPM1 способствует выживанию клетки, будучи связанным с сигнальными путями PI3K/Akt и MAPK/ERK . Количество NPM1 снижается при апоптозе и дифференцировке клеток . Он взаимодействует со многими важными регуляторами апоптоза — белками , и , GAGE и (PI(3,4,5)P3) . После облучения ультрафиолетом NPM1 кратковременно взаимодействует с белком , в результате чего последний теряет способность убиквитинировать p53 и предотвращать апоптоз . NPM1 может взаимодействовать с — активируемой каспазами ДНКазой , которая вносит двуцепочечные разрывы в ДНК и приводит к её в ходе апоптоза — в отсутствие ингибитора этого белка, ICAD , и, таким образом, предотвращает фрагментацию ДНК. Однако антиапоптотическое действие NPM1 зависит от связывания с PI(3,4,5)P3 и АТР: в отсутствие связывания с этими соединениями при апоптозе NPM1 перемещается в нуклеоплазму, где становится нестабильными и впоследствии может быть расщеплён каспазой 3 и разрушен в протеасоме .
В 2015 году было установлено, что NPM1 (а также PARP1) может взаимодействовать с длинной некодирующей РНК Lnc_bc060912, причём через взаимодействие с этими белками Lnc_bc060912 подавляет апоптоз .
Показано, что NPM1 может играть роль в регуляции стабильности и сплайсинга мРНК. Он может выступать в роли PIP3, и комплекс PIP3-NPM1 опосредует антиапоптотическое действие неврального фактора роста (NGF), подавляя ДНКазу, активированную каспазами. Список белков, с которыми взаимодействует NPM1, и соответствующих функций приводится в таблице ниже .
| Процессы и структуры | Белки |
|---|---|
| Репликация, транскрипция, репарация ДНК | MYC; APE1/Ref-1; NFKB1 ; AR ; ; AP2α; HEXIM1; YY1; CBF-A; IRF1; ; GCN5; гистоны; ; ; |
| Контроль клеточного цикла | p53; ARF; MDM2; pRB; p21 ; |
| Биогенез рибосом | EBP1; SENP3 и SENP5; RPL5; RPS9; RPL23; нуклеолин ; p120; NPM3; USP36; ; ; TTF1; FRGY2a/YB1; |
| Репликация вирусов | Rex ( Т-лимфотропный вирус человека ); Rev и Tat (ВИЧ); антигены вируса гепатита дельта ; вируса гепатита В ; p14 ( ); коровые белки вируса гепатита С ; коровый белок вируса японского энцефалита ; основный коровый белок аденовируса; матриксный белок вируса болезни Ньюкасла ; ядерный антиген вируса Эпштейна — Барр |
| Апоптоз | Bax; PARP-1 и PARP-2; PIP3; GAGE |
| Стабильность и сплайсинг мРНК | ; ; NSP 5a3a |
| Модификации, синтез и разрушение белков | ; BRCA1 - ; AKT ; Fbw7γ; HLJ1 |
| Веретено деления, цитоскелет и центромеры | CRM1; RPGR и ; Eg5; ; |
Нокаут Npm1 у мышей приводит к неограниченной дупликации центросом, геномной нестабильности и нарушениям в биогенезе рибосом. Мыши Npm1 −/− характеризуются нарушенным органогенезом ; в частности, передний мозг не развивается должным образом. Такие мыши погибают на эмбриональном этапе в результате анемии , которая является следствием значительных нарушений гемопоэза . Впрочем, мышиные эмбриональные фибробласты , лишённые и p53 , и Npm1 , жизнеспособны и способны к пролиферации в условиях in vitro , следовательно, NPM1 не является белком, строго необходимым для роста и пролиферации клеток. Интересно, что зародыши мышей Npm1 −/− погибают позднее, чем зародыши с утратой функций рибосомных белков, что свидетельствует о важной, но не необходимой роли NPM1 в образовании рибосом .
В рамках иммунной системы NPM1 может выступать в роли молекулярных паттернов, связанных с повреждениями ( англ. damage-associated molecular pattern, DAMP ), или аларминов. Показана роль NPM1 в поддержании жизнеспособности и гематопоэтических стволовых клеток . NPM1 имеет важное значение для функционирования и жизнеспособности зрелых неделящихся нейронов . Впрочем, несмотря на обильную экспрессию NPM1 в мозге , о его конкретных функциях в неделящихся нейронах известно мало .
NPM1 играет важное значение в развитии злокачественных опухолей разных типов, причём он может как стимулировать, так и подавлять рост опухоли. Сверхэкспрессия NPM1 усиливает рост и деление клеток — вероятно, за счёт стимуляции транскрипции рДНК, экспорта рибосомных субъединиц и репликации ДНК в S-фазе. NPM1 может способствовать онкогенезу , мешая работе р53 через ARF. Как правило, уровень NPM1 в опухолевых клетках значительно выше, чем в нормальных. В частности, это характерно для таких опухолей человека, как рак щитовидной железы , глиобластома , гепатоцеллюлярная карцинома , ротовая , , рак толстой кишки , рак яичника , карцинома эндометрия , рак мочевого пузыря и рак простаты . Важную роль в развитии опухоли может играть способность NPM1 к подавлению апоптоза и стимуляции репарации ДНК .
Инактивация гена NPM1 в результате транслокаций или гетерозиготных делеций у человека приводит к злокачественным гематопоэтическим преобразованиям — таким, как острый миелоидный лейкоз (AML), (ALCL), а также к предзлокачественному миелодиспластическому синдрому (MDS). В ходе транслокаций N-концевой домен NPM1 «пришивается» к другим белкам — таким, как киназа , (RARα) и при ALCL, AML и MDS соответственно . При AML мутантная форма NPM1, обозначаемая NPMc+, содержит мутацию в экзоне 12, что приводит к замене остатка триптофана 288 на цистеин. В итоге NPMc+ теряет способность к ядрышковой локализации и перемещениям между ядром и цитоплазмой .
В клетках крови NPM1 выступает как опухолевый супрессор. Это означает, что утрата одной из аллелей NPM1 делает клетки на один шаг ближе к злокачественному перерождению; однако не показано, чтобы NPM1 подавлял гены, активирующие клеточный цикл, индуцировал апоптоз или задержку клеточного цикла при повреждениях ДНК, поэтому его нельзя назвать классическим супрессором опухолей. Скорее его можно назвать зависящим от окружения опухолевым супрессором, то есть ключевое значение в его работе имеют уровень экспрессии, локализация и другие нижестоящие белки, регулирующие клеточный цикл .
Как отмечалось выше, NPM1 чрезвычайно важен для нормального функционирования зрелых нейронов, поэтому он может быть задействован в развитии нейродегенеративных заболеваний .
Показана связь между утратой волос у человека и уровнем экспрессии нуклеофозмина .
Нуклеофозмин может участвовать в развитии некоторых вирусных инфекций . Например, белок капсида вируса некроза нейронов ( англ. Nervous necrosis virus ) связывается с нуклеофозмином в самом начале вирусной инфекции и накапливается в ядре, в частности, в ядрышке. Нокдаун B23 приводил к снижению цитопатических эффектов вируса и угнетению его репликации, поэтому нуклеофозмин способствует репликации вируса и доставляет белок капсида в ядро .