Факторы транскрипции
(транскрипционные факторы) —
белки́
, контролирующие процесс синтеза
мРНК
, а также других видов РНК
на матрице
ДНК
(
транскрипцию
) путём связывания со специфичными участками ДНК
. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (
репрессоры
) или повышение (
активаторы
) константы связывания
РНК-полимеразы
с регуляторными последовательностями регулируемого
гена
.
Определяющая черта факторов транскрипции — наличие в их составе одного или более ДНК-связывающих
доменов
, которые взаимодействуют с характерными участками ДНК, расположенными в регуляторных областях генов. Другие белки, играющие ключевую роль в регуляции
экспрессии генов
, такие как
коактиваторы
,
гистонацетилазы
,
киназы
,
метилазы
, не имеют ДНК-связывающих доменов, и, следовательно, не могут быть причислены к транскрипционным факторам
.
Факторы транскрипции необходимы для регуляции экспрессии генов и обнаружены у всех живых организмов. Их количество, как абсолютное, так и удельное, возрастает с ростом размера
генома
.
В
геноме человека
обнаружено более 2600 белков, имеющих ДНК-связывающий домен, и большинство из них предположительно являются факторами транскрипции
. Следовательно, около 10 % всех генов в геноме кодируют транскрипционные факторы. Таким образом, они являются самым большим семейством белков человека
. Более того, активность многих генов регулируется корпоративным взаимодействием большого числа различных факторов транскрипции, что позволяет обеспечить каждому из генов уникальный способ регуляции в процессе
развития организма
.
Функции
Факторы транскрипции — одна из групп белков, обеспечивающих прочтение и интерпретацию генетической информации. Они связывают ДНК и способствуют инициации программы повышения или понижения транскрипции гена. Таким образом, они жизненно необходимы для нормального функционирования организма на всех уровнях. Ниже перечислены важнейшие из процессов, в которые вовлечены факторы транскрипции.
Регуляция базальной экспрессии генов
Фоновая транскрипционная активность обеспечивается набором ТФ, общим для всех генов. Важный класс
эукариотических
факторов транскрипции — GTFs (general transcription factors)
. Многие из его представителей не связывают ДНК непосредственно, а входят в состав комплекса инициации транскрипции (преинициирующего комплекса), который напрямую взаимодействует с РНК-полимеразой. Наиболее распространенными GTF являются , , (связываются с т. н.
ТАТА-боксом
(элементом
промотора
)), , , и
.
Помимо ТФ, необходимых для экспрессии всех генов, существуют также специфичные факторы транскрипции, обеспечивающие включение/выключение определённых генов в нужный момент.
Многие ТФ многоклеточных организмов вовлечены в обеспечение их развития
. Действуя в соответствии с генетической программой и/или в ответ на внешние воздействия, они инициируют или подавляют транскрипцию определённых генов, что влечет за собой изменения в клеточной морфологии, клеточную дифференциацию,
морфогенез
,
органогенез
и т. д. Например, семейство
гомеобоксных
ТФ критично для формирования правильной морфологии тела у организмов от
дрозофилы
до человека
.
Мутации
генов этих белков (
гомеозисные мутации
) у дрозофил приводят к серьёзным нарушениям в дифференцировке сегментов тела данных насекомых (например, развитие ног вместо усиков).
Другой пример данной группы ТФ — продукт гена полопределяющего региона Y (SRY, Sex-determining Region Y), который играет важную роль в детерминации пола человека.
Ответ на внеклеточные сигналы
Согласованная регуляция взаимодействия клеток многоклеточного организма осуществляется путём высвобождения специальных молекул (
гормонов
,
цитокинов
и т. п.), которые вызывают
сигнальный каскад
в клетках-мишенях. В случае, если сигнал вызывает изменение уровня экспрессии определённых генов, конечным звеном каскада часто оказываются ТФ
.
Эстрогеновый
сигнальный путь — пример короткого каскада, включающего транскрипционный фактор рецептора эстрогена: эстроген секретируется тканями плаценты и яичника, преодолевает
плазматическую мембрану
реципиентных клеток, и связывается со своим рецептором в цитоплазме. Рецептор эстрогена проникает в
ядро
и связывает специфичный участок ДНК, изменяя регуляции транскрипции соответствующего гена
.
Ответ на изменение окружающей среды
ТФ — не единственные конечные звенья сигнальных каскадов, возникающих в ответ на различные внешние стимулы, но они тоже могут быть эффекторами в сигнальных каскадах, индуцируемых воздействием окружающей среды. Например, фактор теплового шока (HSF) активирует гены
белков теплового шока
, которые обеспечивают выживание при повышении температуры (например,
шапероны
)
,
фактор, индуцируемый гипоксией
(HIF) — при снижении концентрации
кислорода
; белок SREBP (sterol regulatory element binding protein) помогает поддерживать необходимое содержание липидов в клетках
.
Контроль клеточного цикла
Многие ТФ, особенно онкогены и онкосупрессоры, участвуют в регуляции
клеточного цикла
. Они определяют переход от одной фазы клеточного цикла к другой, частоту
делений
и интенсивность роста. Один из наиболее известных подобных ТФ — онкоген
Myc
, играющий важную роль в росте клеток и направлении их в
апоптоз
.
Регуляция
Все общебиологические процессы имеют многоуровневую регуляцию и контроль. Это верно и для ТФ — ТФ не только обеспечивают регуляцию уровня накопления белков и РНК в клетке, но и регулируют активность собственных генов (часто с помощью других ТФ). Ниже кратко описаны основные способы регуляции активности ТФ.
Общие для всех белков
Уровень накопления ТФ в клетке регулируется по той же схеме, что и у других белков за счет контроля транскрипции, деградации мРНК,
трансляции
, постпроцессинга белка, его внутриклеточной локализации и деградации. Возможна саморегуляция по принципу
отрицательной обратной связи
— ТФ репрессирует активность кодирующего его гена.
Внутриядерная локализация
У эукариотических организмов процессы транскрипции и трансляции пространственно разделены — они происходят в ядре и
цитоплазме
соответственно. После синтеза ТФ должны проникнуть в ядро, преодолев двойную мембрану. Многие белки, функционирующие в ядре, имеют
сигнал ядерной локализации
— специфичный участок полипептидной цепи, адресующий белок в ядро. Для многих ТФ транслокация является ключевым фактором в регуляции их активности
. Важные классы ТФ, такие как некоторые ядерные рецепторы, должны сперва связать
эндогенный
лиганд
-
агонист
в цитоплазме и только потом транспортироваться в ядро
.
Активация
ТФ могут быть активированы/деактивированны путём воздействия на их сигнал-чувствительный домен различным образом:
связывание лиганда — необходимой для функционирования субстанции, не входящий в состав полипептида (например, ионов Zn
2+
)
фосфорилирование
— многие ТФ должны быть фосфорилированы для получения возможности связывать ДНК.
взаимодействие с другими ТФ и/или корегуляторными белками.
Доступность сайта связывания ДНК
У эукариот гены, не транскрибируемые постоянно, часто находятся в
гетерохроматине
(участках ДНК, плотно упакованных за счет связывания
гистонов
и организованных в компактные хроматиновые фибриллы). ДНК в составе гетерохроматина недоступна для многих факторов транскрипции. Для того, чтобы ТФ могли связаться с ДНК, гетерохроматин должен быть трансформирован в
эухроматин
, обычно путём модификаций гистонов. Также для связывания ТФ с ДНК важную роль играет свобода
хроматина
от
нуклеосом
. Хроматин свободный от нуклеосом называется
открытым хроматином
и значительно чаще связывает факторы транскрипции, чем связанный с нуклеосомами хроматин. Перераспределение нуклеосом осуществляют факторы
ремоделирования хроматина
. Сайт связывания ТФ на ДНК может быть недоступным и в случае, если он связан другим фактором транскрипции. Пары факторов транскрипции могут играть антагонистическую роль (активатор — репрессор) при регуляции активности одного гена.
Наличие других кофакторов/транскрипционных факторов
Большинство ТФ не работают в одиночку. Часто для активации транскрипции гена с его регуляторными элементами должно связаться большое количество ТФ. Связывании ТФ вызывает привлечение промежуточных белков, таких как кофакторы, что приводит к сборке преинициационного комплекса и посадке на промотор РНК-полимеразы.
Структура
ТФ являются модульными по структуре и содержат следующие
домены
:
ДНК-связывающий домен (DBD)
— взаимодействует со специфичными последовательностями ДНК, характерными для
промоторов
и
энхансеров
. Специфичность распознавания определённых последовательностей определяет набор генов, подверженных регуляции данным ТФ;
трансактивирующий домен (TAD)
— содержит участки связывания других белков, например, транскрипционных корегуляторов
;
сигналраспознающий домен (SSD)
(например, лиганд-связывающий домен), который чувствителен к внешнем сигналам и отвечающим за передачу сигнала к другим компонентам транскрипционного комплекса, что вызывает повышение или понижение уровня экспрессии.
ДНК-связывающий домен
Структурно-функциональная единица (домен) факторов транскрипции, связывающая ДНК, называется ДНК-связывающим доменом. Ниже приведен список важнейших семейств ДНК-связывающих доменов/ТФ:
Участки ДНК, которые взаимодействуют с факторами транскрипции, называются сайтами связывания ТФ. Взаимодействие осуществляется за счет
электростатических сил
,
водородных связей
и
сил Ван-дер-Ваальса
. За счет корпоративного, стерически детерминированного действия данных сил, которое определяется пространственной структурой белковой молекулы, ТФ связываться только с определёнными участками ДНК. Не все нуклеотидные основания в ДНК, входящие в сайт связывания ТФ, имеют одинаковую значимость при взаимодействии с белком. Вследствие этого, ТФ обычно связывают не участок со строго определённой первичной структурой, а группу структур с близким сходством, каждую — с разной степенью сродства. Например, хотя консенсусной последовательностью сайта связывания ТАТА-связывающих белков является ТАТАААА, они могут взаимодействовать также с ТАТАТАТ и ТАТАТАА.
Вследствие того, что ТФ взаимодействуют с короткими участками ДНК гетерогенной структуры, потенциальные сайты связывания ТФ могут возникать случайно в достаточно протяженной молекуле ДНК. Маловероятно, однако, что ТФ взаимодействуют со всеми подходящими элементами в геноме.
Различные ограничения, такие как доступность сайтов и наличие кофакторов, могут способствовать направлению ТФ в нужные участки ДНК. Таким образом, затруднительно на основании последовательности генома достоверно предсказать реальное место посадки ТФ на ДНК
in vivo
. Дополнительная специфичность ТФ может опосредоваться наличием нескольких ДНК связывающих доменов в составе одного белка, которые взаимодействуют с двумя или более смежными последовательностями одновременно.
Клинические аспекты
В связи с ключевой ролью ТФ в процессе реализации наследственной информации, некоторые заболевания человека могут быть вызваны мутациями в генах ТФ. Ниже приведены некоторые наиболее изученные нарушения подобного рода:
Диабеты
. Редкая форма
диабета
, называемая
MODY
(Maturity onset diabetes of the young) может быть обусловлена мутациями в генах некоторых ТФ
.
Developmental verbal dyspraxia
. (нарушение речевых функций). Мутации в гене ТФ
FOXP2
ассоциированы с развитием данного заболевания, при котором человек не может производить координированных движений, необходимых для речевой функции
.
Аутоиммунные заболевания
. Мутации в гене ТФ
FOXP3
связаны с аутоиммунным заболеванием IPEX (immune dysregulation polyendocrinopathy enteropathy X-linked syndrome)
.
Рак
. Многие факторы транскрипции являются онкогенами или онкосупрессорами, и их мутации или неправильная регуляция могут приводить к развитию рака. Например, синдром Li-Fraumeni обусловлен мутациями в гене онкосупрессора
p53
.
Классификация
ТФ могут классифицироваться по (1) механизму действия, (2) регуляторной функции, (3) структуре ДНК-связывающего домена, а также на натуральные и (5)искусственные.
Механизм действия
По данному признаку выделяют три класса ТФ:
Главные факторы транскрипции
(GTFs)
, вовлеченные в образование инициационного комплекса. Наиболее важные из них — TFIIA, TFIIB, TFIID, TFIIE, TFIIF, и TFIIH. Они присутствуют во всех клетках и взаимодействуют с кором промотора генов, транскрибируемых РНК-полимеразой второго класса.
ТФ, взаимодействующие с upstream-участками ДНК
, (областями, расположенными до промотора, лежащими относительно него с другой стороны от
кодирующей области гена
).
Индуцируемые ТФ
сходны с предыдущим классом, но требуют активации либо ингибирования.
Функция
Конститутивные
— присутствуют всегда во всех клетках — главные факторы транскрипции,
Sp1
,
NF1
, .
Активируемые
(активны в определённых условиях)
Участвующие в развитии организма
(клетко-специфичные) — экспрессия строго контролируется, но, начав экспрессироваться, не требуют дополнительной активации — GATA, HNF, PIT-1, MyoD, Myf5, Hox, Winged Helix.
Сигнал-зависимые
— требуют внешнего сигнала для активации
резидентные ядерные факторы
— находятся в ядре независимо от активации — CREB, AP-1, Mef2
латентные цитоплазматические факторы
— в неактивном состоянии локализованы в цитоплазме, после активации транспортируются в ядро — STAT, R-SMAD,
NF-kB
,
Notch
, TUBBY, NFAT.
Структурная классификация
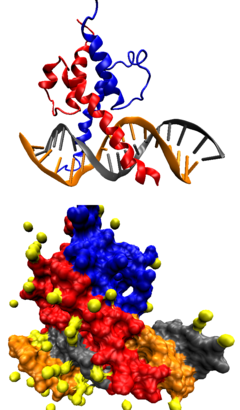
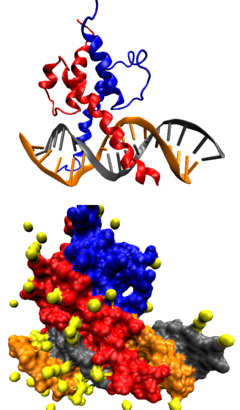
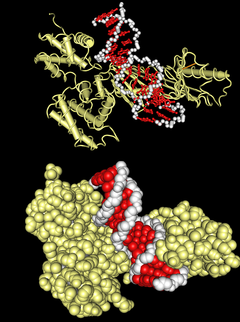
ДНК-связывающий домен типа «
лейциновая молния
» в комплексе с ДНК. Вверху — схематичное изображение молекулярной поверхности, внизу — третичной структуры комплекса.
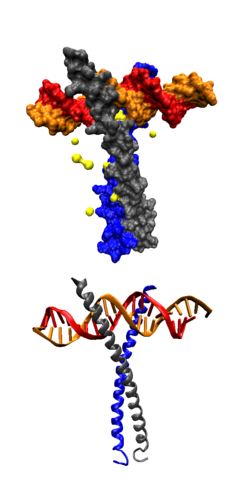
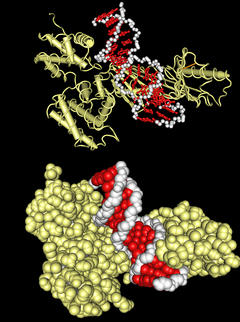
ДНК-связывающий домен типа «
спираль-петля-спираль
» в комплексе с ДНК. Вверху — схематичное изображение третичной структуры, внизу — молекулярной поверхности комплекса.
Факторы транскрипции классифицируют на основании сходства первичной структуры (что предполагает и сходство третичной структуры) ДНК-связывающих доменов
.
Систему
CRISPR
можно адаптировать так, чтобы она действовала как транскрипционный фактор (crisprTF). Для этого CRISPR-ассоциированный белок, известный как
Cas9
, изменяют так, чтобы он после связывания с ДНК больше не мог её расщепить. Затем к нему добавляют сегмент, который активирует или подавляет экспрессию генов путём модуляции транскрипционного механизма клетки
. В отличие от транскрипционных факторов на базе
цинковых пальцев
и
, для узнавания ДНК системе CRISPR-Cas требуется только создание соответствующей последовательности РНК-«гида», а не создание новых белковых доменов фермента, что делает его гораздо более доступным благодаря дешевизне и простоте (вплоть до того что разработан набор правил — «грамматика» — описывающих, как спроектировать синтетический транскрипционный фактор (STFS) и программа для его автоматизированного проектирования
).
Karin M.
Too many transcription factors: positive and negative interactions
(англ.)
// New Biol. : journal. — 1990. — Vol. 2 , no. 2 . — P. 126—131 . — .
Roeder R.G.
The role of general initiation factors in transcription by RNA polymerase II
(англ.)
//
(англ.)
(: journal. — 1996. — Vol. 21 , no. 9 . — P. 327—335 . —
doi
: . — .
Lee T.I., Young R.A.
Transcription of eukaryotic protein-coding genes
(англ.)
//
Annu. Rev. Genet.
: journal. — 2000. — Vol. 34 . — P. 77—137 . —
doi
: . — .
Mitchell P.J., Tjian R.
Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins
(англ.)
// Science : journal. — 1989. — Vol. 245 , no. 4916 . — P. 371—378 . —
doi
: . — .
Ptashne M., Gann A.
Transcriptional activation by recruitment
(англ.)
// Nature. — 1997. — Vol. 386 , no. 6625 . — P. 569—577 . —
doi
: . — .
↑
Brivanlou A.H., Darnell J.E.
Signal transduction and the control of gene expression
(англ.)
// Science : journal. — 2002. — Vol. 295 , no. 5556 . — P. 813—818 . —
doi
: . — .
van Nimwegen E.
Scaling laws in the functional content of genomes
(англ.)
//
(англ.)
(: journal. — 2003. — Vol. 19 , no. 9 . — P. 479—484 . —
doi
: . — .
Babu M.M., Luscombe N.M., Aravind L., Gerstein M., Teichmann S.A.
Structure and evolution of transcriptional regulatory networks
(англ.)
// Curr. Opin. Struct. Biol. : journal. — 2004. — Vol. 14 , no. 3 . — P. 283—291 . —
doi
: . — .
Lambert S. A. , Jolma A. , Campitelli L. F. , Das P. K. , Yin Y. , Albu M. , Chen X. , Taipale J. , Hughes T. R. , Weirauch M. T.
(англ.)
// Cell. — 2018. — 8 February (vol. 172 , no. 4). — P. 650—665 . —
doi
: . — . [ ]
Reese J.C.
Basal transcription factors
(неопр.)
// Current opinion in genetics & development. — 2003. — April (т. 13 , № 2). — С. 114—118 . —
doi
: . — .
Shilatifard A., Conaway R.C., Conaway J.W.
The RNA polymerase II elongation complex
(англ.)
//
(англ.)
(: journal. — 2003. — Vol. 72 . — P. 693—715 . —
doi
: . — .
Thomas M.C., Chiang C.M.
The general transcription machinery and general cofactors
(англ.)
// Critical reviews in biochemistry and molecular biology : journal. — 2006. — Vol. 41 , no. 3 . — P. 105—178 . — .
Lobe C.G.
Transcription factors and mammalian development
(неопр.)
// Current topics in developmental biology. — 1992. — Т. 27 . — С. 351—383 . — .
Lemons D., McGinnis W.
Genomic evolution of Hox gene clusters
(англ.)
//
Science
: journal. — 2006. — September (vol. 313 , no. 5795). — P. 1918—1922 . —
doi
: . — .
Moens C.B., Selleri L.
Hox cofactors in vertebrate development
(неопр.)
// Developmental biology. — 2006. — March (т. 291 , № 2). — С. 193—206 . —
doi
: . — .
Ottolenghi C., Uda M., Crisponi L., Omari S., Cao A., Forabosco A., Schlessinger D.
Determination and stability of sex
(неопр.)
// BioEssays : news and reviews in molecular, cellular and developmental biology. — 2007. — January (т. 29 , № 1). — С. 15—25 . —
doi
: . — .
Pawson T.
Signal transduction--a conserved pathway from the membrane to the nucleus
(англ.)
// Developmental genetics : journal. — 1993. — Vol. 14 , no. 5 . — P. 333—338 . —
doi
: . — .
Osborne C.K., Schiff R., Fuqua S.A., Shou J.
Estrogen receptor: current understanding of its activation and modulation
(англ.)
//
(англ.)
(: journal. — 2001. — December (vol. 7 , no. 12 Suppl). — P. 4338s—4342s; discussion 4411s—4412s . — .
Shamovsky I., Nudler E.
(англ.)
//
Cell. Mol. Life Sci.
: journal. — 2008. — March (vol. 65 , no. 6). — P. 855—861 . —
doi
: . — .
Benizri E., Ginouvès A., Berra E.
The magic of the hypoxia-signaling cascade
(англ.)
//
Cell. Mol. Life Sci.
: journal. — 2008. — April (vol. 65 , no. 7—8). — P. 1133—1149 . —
doi
: . — .
Weber L.W., Boll M., Stampfl A.
(англ.)
//
(англ.)
(: journal. — 2004. — November (vol. 10 , no. 21). — P. 3081—3087 . — .
11 августа 2007 года.
↑
Whiteside S.T., Goodbourn S.
(англ.)
//
(англ.)
(: journal. —
(англ.)
(, 1993. — April (vol. 104 (Pt 4)). — P. 949—955 . — .
Bohmann D.
Transcription factor phosphorylation: a link between signal transduction and the regulation of gene expression
(англ.)
// Cancer cells (Cold Spring Harbor, N.Y. : 1989) : journal. — 1990. — November (vol. 2 , no. 11). — P. 337—344 . — .
Weigel N.L., Moore N.L.
Steroid Receptor Phosphorylation: A Key Modulator of Multiple Receptor Functions
(англ.)
: journal. — 2007. — .
Wärnmark A., Treuter E., Wright A.P., Gustafsson J-Å.
Activation functions 1 and 2 of nuclear receptors: molecular strategies for transcriptional activation
(англ.)
//
(англ.)
(: journal. — 2003. — Vol. 17 , no. 10 . — P. 1901—1909 . —
doi
: . — .
Littlewood T.D., Evan G.I.
Transcription factors 2: helix-loop-helix
(неопр.)
// Protein profile. — 1995. — Т. 2 , № 6 . — С. 621—702 . — .
Vinson C., Myakishev M., Acharya A., Mir A.A., Moll J.R., Bonovich M.
Classification of human B-ZIP proteins based on dimerization properties
(англ.)
//
(англ.)
(: journal. — 2002. — September (vol. 22 , no. 18). — P. 6321—6335 . —
doi
: . — . —
PMC
.
Wintjens R., Rooman M.
Structural classification of HTH DNA-binding domains and protein-DNA interaction modes
(англ.)
//
(англ.)
(: journal. — 1996. — September (vol. 262 , no. 2). — P. 294—313 . —
doi
: . — .
Gehring W.J., Affolter M., Bürglin T.
Homeodomain proteins
(англ.)
//
(англ.)
(: journal. — 1994. — Vol. 63 . — P. 487—526 . —
doi
: . — .
Dahl E., Koseki H., Balling R.
Pax genes and organogenesis
(неопр.)
// BioEssays : news and reviews in molecular, cellular and developmental biology. — 1997. — September (т. 19 , № 9). — С. 755—765 . —
doi
: . — .
Laity J.H., Lee B.M., Wright P.E.
Zinc finger proteins: new insights into structural and functional diversity
(англ.)
// Current opinion in structural biology : journal. — 2001. — February (vol. 11 , no. 1). — P. 39—46 . —
doi
: . — .
Fichou Y., Nectoux J., Bahi-Buisson N., Rosas-Vargas H., Girard B., Chelly J., Bienvenu T.
The first missense mutation causing Rett syndrome specifically affecting the MeCP2_e1 isoform.
(англ.)
// Neurogenetics : journal. — 2008. — November. — .
Al-Quobaili F., Montenarh M.
Pancreatic duodenal homeobox factor-1 and diabetes mellitus type 2 (review).
(англ.)
//
(англ.)
(: journal. — 2008. — Vol. 21(4) . — P. 399—404 . — .
Lai C.S., Fisher S.E., Hurst J.A., Vargha-Khadem F., Monaco AP.
A forkhead-domain gene is mutated in a severe speech and language disorder.
(англ.)
// Nature : journal. — 2001. — Vol. 413(6855) . — P. 519—523 . — .
↑
Banerjee-Basu S., Baxevanis A.D.
Structural analysis of disease-causing mutations in the P-subfamily of forkhead transcription factors.
(англ.)
// Proteins : journal. — 2004. — Vol. 54(4) . — P. 639—647 . — .
Ariffin H., Martel-Planche G., Daud S.S., Ibrahim K., Hainaut P.
Li-Fraumeni syndrome in a Malaysian kindred.
(неопр.)
// Cancer Genet Cytogenet.. — 2008. — Т. 186(1) . — С. 49—53 . — .
Stegmaier P., Kel A.E., Wingender E.
(англ.)
// Genome informatics. International Conference on Genome Informatics : journal. — 2004. — Vol. 15 , no. 2 . — P. 276—286 . — .
19 июня 2013 года.
Matys V., Kel-Margoulis O.V., Fricke E., Liebich I., Land S., Barre-Dirrie A., Reuter I., Chekmenev D., Krull M., Hornischer K., Voss N., Stegmaier P., Lewicki-Potapov B., Saxel H., Kel A.E., Wingender E.
TRANSFAC and its module TRANSCompel: transcriptional gene regulation in eukaryotes
(англ.)
//
(англ.)
(: journal. — 2006. — Vol. 34 , no. Database issue . — P. D108—10 . —
doi
: . — .
(неопр.)
.
Дата обращения: 5 августа 2007.
21 марта 2012 года.
Qi Lei S. , Larson Matthew H. , Gilbert Luke A. , Doudna Jennifer A. , Weissman Jonathan S. , Arkin Adam P. , Lim Wendell A.
(англ.)
// Cell. — 2013. — February (vol. 152 , no. 5). — P. 1173—1183 . —
ISSN
. —
doi
: . — . [ ]
Farzadfard Fahim , Perli Samuel D. , Lu Timothy K.
(англ.)
// ACS Synthetic Biology. — 2013. — 11 September (vol. 2 , no. 10). — P. 604—613 . —
ISSN
. —
doi
: . — . [ ]
Gilbert Luke A. , Larson Matthew H. , Morsut Leonardo , Liu Zairan , Brar Gloria A. , Torres Sandra E. , Stern-Ginossar Noam , Brandman Onn , Whitehead Evan H. , Doudna Jennifer A. , Lim Wendell A. , Weissman Jonathan S. , Qi Lei S.
// Cell. — 2013. — Июль (т. 154 , № 2). — С. 442—451 . —
ISSN
. —
doi
: . — . [ ]
Perez-Pinera Pablo , Kocak D Dewran , Vockley Christopher M , Adler Andrew F , Kabadi Ami M , Polstein Lauren R , Thakore Pratiksha I , Glass Katherine A , Ousterout David G , Leong Kam W , Guilak Farshid , Crawford Gregory E , Reddy Timothy E , Gersbach Charles A.
(англ.)
// Nature Methods. — 2013. — 25 July (vol. 10 , no. 10). — P. 973—976 . —
ISSN
. —
doi
: . — . [ ]
Purcell Oliver , Peccoud Jean , Lu Timothy K.
(англ.)
// ACS Synthetic Biology. — 2014. — 3 January (vol. 3 , no. 10). — P. 737—744 . —
ISSN
. —
doi
: . — . [ ]