Водоросли
- 1 year ago
- 0
- 0

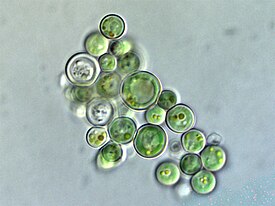
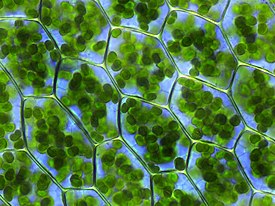
Глаукофи́товые во́доросли , или глаукофи́ты , или глаукоцистофи́ты ( лат. Glaucophyta ) — небольшой древний отдел одноклеточных водорослей, включающий восемь родов и 21 вид . Глаукофиты особенно интересны своими уникальными хлоропластами ( цианеллами ), довольно примитивно устроенными по сравнению с хлоропластами других водорослей: они содержат слой муреина между мембранами и характеризуются множеством других свойств, присущих цианобактериям . Согласно современным представлениям, глаукофитовые водоросли обособились в отдельную группу у самого основания Archaeplastida .
Название группы образовано от др.-греч. γλαυκός — «сине-зелёный» и φυτόν — «растение» и обусловлено характерным сине-зелёным цветом их хлоропластов .
Изучение глаукофитов восходит к 1854 году , когда немецкий врач и ботаник Герман Ицигсон (1814—1879) обнаружил в сфагновых болотах близ Нойдамма (совр. Дембно , Польша) одноклеточную водоросль c коккоидным типом организации, отнесённую к новому роду глаукоцистис ( Glaucocystis ) и виду Glaucocystis nostochinearum . Краткие описания рода и вида были опубликованы в 1868 году — в добавлениях к последнему тому изданного Людвигом Рабенхорстом трёхтомного энциклопедического труда о водорослях «Flora Europaea. Algarum aquae dulcis et submarinae». Несмотря на то, что в описаниях отмечалось наличие хорошо различимого ядра , род Glaucocystis был тогда отнесён к сине-зелёным водорослям и включён в семейство ; в 1930-х — 1950-х годах (а иногда и позднее) альгологи относили род к зелёным водорослям , включая его в семейство .
В 1883 году шведский ботаник Густаф Лагерхейм описал, изучив собранные им в 1882 году в окрестностях Уппсалы образцы водорослей, новые род глеохете ( Gloeochaete ) и вид Gloeochaete wittrockiana — уже с пальмеллоидным типом организации; этот род он отнёс к уже упоминавшемуся семейству . В 1924 году советский альголог А. А. Коршиков при изучении проб, взятых на Харьковщине из грязных луж, обнаружил своеобразного фототрофного жгутиконосца , которого описал в своей статье как род цианофора ( Cyanophora ) и вид Cyanophora paradoxa ; эту водоросль длительное время относили к криптофитам . Позднее были описаны и другие роды и виды глаукофитов .
В первой трети XX века в трудах ряда альгологов, и прежде всего в работах немецкого учёного Адольфа Пашера , оформилась точка зрения, по которой глаукофиты представляют собой пример внутриклеточного симбиоза , когда в цитоплазме эукариотического организма- хозяина поселяются симбионты - цианобактерии . Пашер назвал такой тип симбиоза синцианозом , хозяина — цианомом , симбионтов — цианеллами ; только в конце XX века изучение ультраструктуры и геномов цианелл глаукофитов показало, что они хотя и произошли некогда от цианобактерий, но не являются отдельными организмами, а представляют собой своеобразную разновидность хлоропластов (иногда такие хлоропласты именуют муропластами ) .
При этом глаукофитов продолжали трактовать как сборную группу водорослей. Только в 1954 году латвийский альголог Генрих Скуя выделил их в самостоятельный отдел Glaucophyta , предпочитая рассматривать синцианозы как «составные организмы», характеризующиеся появлением новых признаков, которыми не обладали по отдельности партнёры по симбиозу . В 1986 году немецкие ботаники Людвиг Кис и Бруно Кремер предложили альтернативное, типифицированное название таксона: Glaucocystophyta . Монофилия таксона была убедительно обоснована проведёнными в конце XX — начале XXI века молекулярно-филогенетическими исследованиями .
В 1981 году английский биолог Томас Кавалье-Смит предложил новую трактовку царства Plantae (которое в традиционном, геккелевском понимании представляло собой полифилетическую группу), включив в него только глаукофитов, красные водоросли и зелёные растения (т. e. те группы фотосинтезирующих организмов, хлоропласты которых возникли в результате первичного эндосимбиоза — поглощения эукариотным организмом путём фагоцитоза свободноживущих цианобактерий, которые после горизонтального переноса генов от симбионта в ядро хозяина эволюционировали в двумембранные хлоропласты ). Одновременно Кавалье-Смит объединил глаукофитовые и красные водоросли в новый таксон , включив в диагноз последнего следующие общие для этих двух отделов признаки: наличие фико били сом , одиночные (не сгруппированные в граны ) тилакоиды , откладывание зёрен крахмала в цитозоле (а не в пластидах). При этом в качестве характерного признака глаукофитов, отличающего их от красных водорослей, было указано наличие пептидогликана в оболочке хлоропластов .
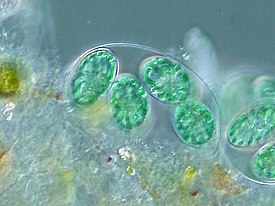
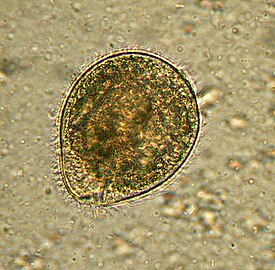
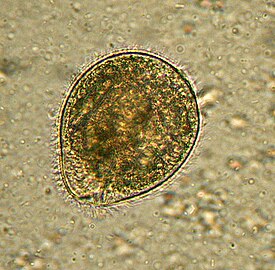
Глаукофитовые водоросли представлены одноклеточными и колониальными формами с монадным, коккоидным и пальмеллоидным типами организации . Для всех глаукофитов характерны эллипсоидальная форма клетки и микроскопические размеры. При этом у водорослей с пальмеллоидным строением таллома клетки имеют почти сферическую форму диаметром 10—15 мкм у Gloeochaete и 24–36 мкм у Cyanoptyche ; у видов рода Glaucocystis , представляющих коккоидный тип организации, клетки сильно вытянуты, причём их размеры у разных видов варьируют: у G. nostochinearum клетки имеют 15—24 мкм в длину и 7,5—13,5 мкм в поперечнике, а у G. reniformis — 25—32 мкм в длину и 12—30 мкм в поперечнике .
Виды рода Cyanophora характеризуются монадным строением таллома и достаточно чётко подразделяются на две группы. В одну входят виды с овоидной или эллипсоидальной формой клетки: C. paradoxa (примерно 15 мкм в длину и 9 мкм в поперечнике ), C. cuspidata , C. kugrensii ; в другую — виды с клетками бобовидной формы, уплощёнными в дорсовентральном направлении: C. biloba (10—15 мкм в длину, 6—9 мкм в ширину и 3—4 мкм в толщину ), C. sudae , C. tetracyana .
Под клеточной мембраной находятся уплощённые везикулы (альвеолы), причём у некоторых форм они содержат чешуйкоподобные структуры, у других — фибриллярный материал, а у некоторых они пустые . Неподвижные вегетативные клетки колониальных глаукофитов имеют прочные клеточные стенки , которые, как правило, состоят из целлюлозы ; в то же время у подвижных жгутиконосцев из рода Cyanophora клеточные стенки отсутствуют . Таким образом, покровы клеток глаукофитовых водорослей напоминают таковые у Alveolata , однако гомологии здесь нет .
Если у представителей Cyanophora уплощённые везикулы полностью покрывают протопласт , частично перекрывая друг друга и образуя гребни на поверхности клеток, то у видов Glaucocystis везикулы, напротив, более или менее отделены друг от друга зазорами. Исследователи предполагают, что утрата перекрытия везикул у предков рода Glaucocystis была связана с формированием у них прочной клеточной стенки, расположенной снаружи от цитоплазматической мембраны и обеспечивающей более эффективную защиту протопласта . Клеточная стенка глаукоцистиса имеет в толщину 1,5—2,5 мкм ; при этом она чрезвычайно сходна по размерам и морфологии с клеточной стенкой коккоидных зелёных водорослей рода , но отличается чрезвычайно высоким (до 90 %) содержанием α-целлюлозы .
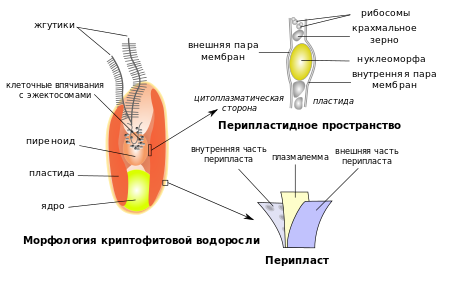
Подвижные клетки глаукофитов (монадные формы и стадии жизненного цикла) имеют по два жгутика неодинаковой длины (передний жгутик короче, задний — длиннее). Строение жгутиков типично для эукариот : аксонема жгутика образована девятью парами микротрубочек на периферии и двумя микротрубочками в центре . При этом выходы внешних частей жгутиков за пределы клетки располагаются в диаметрально противоположных местах её поверхности ; это отличает жгутики глаукофитов от жгутиков криптофитовых водорослей , у которых оба жгутика отходят от возвышения на переднем конце клетки .
Каждый жгутик несёт два ряда тонких фибриллярных (нетрубчатых) волосков, схожих с волосками на жгутиках некоторых зелёных водорослей , но, однако, отличных от трубчатых мастигонем охрофитовых и криптофитовых водорослей . У некоторых представителей глаукофитов корешковая система жгутика организована подобно тому, как это имеет место у некоторых зелёных водорослей (микротрубочковые корешки расположены крестообразно); у других представителей имеются многослойные структуры, как у харовых водорослей .
Происхождение глаукофитов с немонадным типом организации от жгутиконосных предков достаточно очевидно. Так, от клеток Gloeochaete отходят по два длинных волоска-псевдожгутика, отличающихся от настоящих жгутиков отсутствием центральной пары микротрубочек. У клеток Glaucocystis сохраняются два рудиментарных жгутика, которые за пределы клеточной стенки не выходят .
Митохондрии глаукофитов имеют пластинчатые кристы на своей внутренней мембране ; в этом они сходны с зелёными и красными , а также с криптофитовыми водорослями . Митохондриальные геномы глаукофитов в целом сходны с таковыми у других Archaeplastida, они не содержат необычных генов и не имеют других особенностей генома. Состав генов мтДНК консервативен среди различных видов и сравним с таковым у наиболее богатых генами мтДНК зелёных и красных водорослей .
В ходе исследований митохондриальных геномов глаукофитов у некоторых представителей таксона были выявлены отклонения от стандартного генетического кода , причём могут использоваться различные неканонические старт- и стоп-кодоны .
Глаукофитовые водоросли привлекают внимание исследователей благодаря своим уникальным хлоропластам , которые являются особой древней ветвью эволюции пластид . Их называют цианеллами , или муропластами . Хлоропласты глаукофитов имеют округлую или эллипсоидную форму. Они окружены двумя , между которыми расположен слой муреина (пептидогликана), причём его толщина меньше, чем в среднем у бактерий . Из-за наличия муреина пластиды глаукофитов чувствительны к β-лактамным антибиотикам , а их клеточная стенка лизируется лизоцимом . Пептидогликан глаукофитов характеризуется наличием N-ацетил путресцина в качестве заместителя в половине 1- карбоксигрупп остатков глутаминовой кислоты в пептидных цепочках. Биосинтез предшественников пептидогликана глаукофитов происходит в строме, а сборка полимерной сети происходит в периплазматическом пространстве. Интересно, что большая часть ферментов , участвующих в биосинтезе пептидогликана, у Cyanophora paradoxa кодируется не пластидным, а ядерным геномом и импортируется в пластиды .
Хлоропласты имеют сине-зелёный цвет, поскольку зелёный хлорофилл a маскируется синими пигментами фикоцианином и аллофикоцианином . Подобно цианобактериям и красным водорослям , у глаукофитов эти пигменты находятся в фикобилисомах , расположенных на поверхности тилакоидов . При этом фикоэритрин (наиболее обильно представленный пигмент в пластидах красных водорослей) и типичные для цианобактерий каротиноиды — и — отсутствуют. От фикобилисом красных водорослей фикобилисомы глаукофитов отличаются и по некоторым структурным особенностям . Каротиноиды представлены β-каротином , зеаксантином и . Некоторые пигменты ( феофорбид a , β-криптоксантин и зеаксантин) глаукофитовой водоросли оказывают антипролиферативное действие на клетки рака молочной железы , лёгких и меланомы .
Тилакоиды не собраны в граны, располагаются одиночно и равноудалённо друг от друга, как у цианобактерий и красных водорослей. В центре пластид располагаются крупные одиночные или мелкие многочисленные тельца, похожие на карбоксисомы . Они содержат фермент Рубиско и принимают участие в концентрировании углекислого газа (в хлоропластах эукариот эту функцию обычно выполняет пиреноид ). Карбоксисомо-подобные тельца Cyanophora не имеют белковой оболочки, как карбоксисомы бактерий, но и не пронизываются тилакоидом, как пиреноид в хлоропластах прочих эукариот. Кроме того, в пластидах имеются полифосфатные гранулы . Наличие муреина и карбоксисомо-подобных телец — плезиоморфные признаки Archaeplastida, которые глаукофиты сохранили. Интересно, что амёба из супергруппы Rhizaria также имеет фотосинтетические органеллы со слоем муреина, карбоксисомами и концентрическими тилакоидами, которые по строению напоминают пластиды глаукофитов. Тем не менее, данные филогенетического анализа убедительно свидетельствуют о независимом происхождении этих двух фотосинтетических органелл .
Была предложена гипотеза, связывающая наличие муреина и карбоксисомо-подобных телец в хлоропластах глаукофитов. Согласно этой гипотезе, если карбоксисомо-подобные тельца действительно выполняют роль карбоксисом в концентрировании СО 2 , то внутри пластиды должна быть высокая концентрация неорганического иона HCO 3 - . Высокая концентрация данного иона может создавать заметное различие между осмолярностью внутри пластиды и в цитозоле , что угрожает целостности мембраны органеллы. Поэтому наличие прочной муреиновой стенки в пластидах глаукофитов может служить удобным и малозатратным по энергии механизмом поддержания целостности органеллы .
Как и у цианобактерий, хлоропластная ДНК сконцентрирована в центре хлоропласта. Геном хлоропластов глаукофитовых водорослей содержит ряд , типичных для цианобактерий, и кодирует некоторые белки , которые геномы хлоропластов других водорослей не кодируют. В частности, в нём имеется ген транспортно-матричной РНК ( тмРНК ) — особой малой РНК , характерной для бактерий, а также пластид и митохондрий примитивных организмов . Более того, в нём закодированы обе субъединицы Рубиско, в то время как у высших растений, эвгленовых и зелёных водорослей этот ген перенёсся в ядерный геном . Некоторые особенности транспорта белков в хлоропластах глаукофитов также сближают их с цианобактериями .
Длительное время их даже рассматривали как эндосимбиотические синезелёные водоросли, живущие в одноклеточных гетеротрофных эукариотах. Их также рассматривали как переходную стадию между эндосимбиотическими цианобактериями и настоящими пластидами. Тем не менее, геном хлоропластов глаукофитовых водорослей примерно в 10 раз меньше генома свободноживущих цианобактерий и по размерам (около 140000 пар нуклеотидов ) близок к геному хлоропластов других водорослей. В отличие от цианобактерий, хлоропласты глаукофитов не кодируют белков дыхательной ЭТЦ . Механизм деления хлоропластов у глаукофитовой водоросли Cyanophora paradoxa занимает среднее положение между механизмом деления клеток цианобактерий и пластид. У пластид в районе деления формируются внутреннее и внешнее кольца электрон-плотного материала. У хлоропластов Cyanophora paradoxa имеется лишь внутреннее кольцо . В делении хлоропластов у этой водоросли, как и у бактерий, ключевую роль играет белок .
Как и пластиды других водорослей, хлоропласты глаукофитов не могут размножаться вне клетки и не могут культивироваться вне цитоплазмы клетки-хозяина, хотя в изолированном виде сохраняют структурную целостность гораздо дольше, чем у хлоропластов высших растений (однако скорости фиксации углекислого газа и выделения кислорода у них снижаются намного быстрее). На основании этих данных большинство исследователей считают хлоропласты глаукофитовых водорослей органеллами (примитивными пластидами), а не эндосимбиотическими цианобактериями .
Фитохромы Cyanophora и Gloeochaete по фотосенсорным характеристикам не имеют подобных себе среди других эукариотических фитохромов. В частности, они чувствительны к синему свету, как многие фитохромы цианобактерий. Фотоциклы (то есть конформационные изменения молекулы хромофора в зависимости от длины волны поглощённого света) у Cyanophora и Gloeochaete несколько различаются: для первой водоросли характерен сине-дальнекрасный фотоцикл, а для второй — красно-синий. Таким образом, видимо, фотосенсорные характеристики фитохромов глаукофитов варьируют. В настоящее время изучается физиологическая роль необычных фитохромов глаукофитов .
Запасной продукт в клетках глаукофитовых водорослей — крахмал , откладывается в виде гранул в цитозоле . В этом отношении глаукофиты схожи с красными водорослями и отличаются от Viridiplantae , которые накапливают запасные вещества в пластидах. Однако красные водоросли запасают не обычный крахмал, как глаукофиты и Viridiplantae, а особое амилопектин -подобное соединение .
Размножение у глаукофитов — вегетативное и бесполое (у Glaucocystis — с помощью , у Gloeochaete и Cyanoptyche — с помощью зооспор ; Cyanophora обычно размножается продольным делением пополам, но может также образовывать зооспоры). Половое размножение не описано. Митоз открытый, цитокинез продольный .
Глаукофитовые водоросли обитают исключительно в пресных водах, в том числе в болотах . Крупные популяции глаукофитов встречаются редко . Среди них есть и планктонные формы, и прикреплённые колонии. В пробах глаукофиты выделяются от других водорослей со схожей морфологией по наличию пластид яркого сине-зелёного цвета. От синезелёных водорослей их отличает наличие неокрашенной цитоплазмы .
Глаукофитовые водоросли — чрезвычайно древняя группа; их даже называют « целакантами эндосимбиоза». Некоторые исследователи предполагают, что в рамках группировки Archaeplastida ( дивергенция которой произошла, по имеющимся оценкам, примерно 1,5—1,6 млрд лет тому назад ) глаукофиты выделились в самостоятельную группу ещё до расхождения зелёных и красных водорослей . Филогенетический анализ ядерных генов рРНК свидетельствует о монофилии глаукофитов .
Анализ генов рРНК хлоропластов также говорит о монофилии хлоропластов глаукофитов, несмотря на разнообразие размеров, форм и ультраструктуры. Судя по всему, хлоропласты глаукофитов — это архаичная ветвь, которая после своего обособления не получила дальнейшего развития. О примитивности хлоропластов глаукофитов свидетельствует наличие у них только хлорофилла a и отсутствие вспомогательных хлорофиллов, присутствие фикобилисом, одиночных и равноудалённых тилакоидов, карбоксисом, слоя муреина, а также характерных для цианобактерий последовательностей в геноме хлоропластов. Таким образом, по современным представлениям хлоропласты глаукофитов в эволюционном плане занимают промежуточное положение между цианобактериями и хлоропластами других водорослей .
Данные о положении глаукофитов среди эукариот достаточно противоречивы; анализ филогении осложняется тем, что скорость эволюции генома у глаукофитов (как, впрочем, и у катаблефарид — одного из отрядов в типе Cryptista ) существенно ниже, чем в других ветвях эукариот . По-видимому, этим объясняются результаты выполненного японскими биологами в 2009 году филогенетического исследования, согласно которому глаукофиты и криптофиты являются сестринскими группами , хотя они не имеют очевидного сходства по морфологическим и ультраструктурным характеристикам; при этом глаукофиты с криптофитами и красные водоросли — базальные ветви по отношению к кладе , состоящей из зелёных растений и (так что в этом исследовании монофилия Archaeplastida не нашла подтверждения) .
Между тем в том же 2009 году коллектив исследователей из разных стран провёл масштабное филогенетическое исследование, в ходе которого анализировались полученные от 72 видов эукариот последовательности ядерной ДНК (127 генов); результаты анализа подтвердили монофилию Archaeplastida . Аналогичными оказались и результаты последующих исследований: по 124 генам рибосомальной ДНК (2012) , 258 генам ядерной ДНК (2012) , 157 генам ядерной ДНК (2014) , 14 генам митохондриальной ДНК (2014) и др. В настоящее время монофилия Archaeplastida может считаться достаточно твёрдо установленной; в то же время порядок ветвления трёх групп (глаукофиты, красные водоросли, зелёные растения), составляющих данную кладу, остаётся неясным: результаты различных исследований противоречат друг другу .
В макросистеме живущих организмов, предложенной в 2015 году Майклом Руджеро с соавторами, отдел Glaucophyta Skuja , 1954 (включающий единственный класс Glaucophyceae с единственным порядком Glaucocystales ) вместе с отделом Rhodophyta (красные водоросли) составляет подцарство Cavalier-Smith , 1981 царства Plantae Haeckel , 1866 em. Cavalier-Smith , 1981 (= Archaeplastida Adl et al. , 2005 ); другое подцарство — Viridiplantae — включает зелёные растения. При этом в зависимости от порядка ветвления трёх перечисленных групп подцарство Biliphyta представляет собой либо монофилетический (если сестринской группой для глаукофитов являются красные водоросли ), либо парафилетический (если в роли сестринской группы выступают зелёные растения или если ветвь глаукофитов выделилась до расхождения двух других групп ) таксон .
В статье 1986 года, посвящённой вопросам классификации глаукофитов, Л. Кис и Б. П. Кремер выделили 9 родов и около 13 видов глаукофитовых водорослей . Эти же цифры фигурировали и в ряде последующих публикаций ; между тем классификация глаукофитов постепенно изменялась, и в глобальной базе данных AlgaeBase ?! , содержащей информацию по всем группам водорослей и морских трав , к ноябрю 2015 года упоминались 8 родов и 21 вид глаукофитов (с учётом монотипического рода Cyanoptyche , отнесённого в базе данных к зелёным растениям — к семейству порядка Chlamydomonadales класса Chlorophyceae , в то время как молекулярные данные однозначно указывают на его принадлежность к глаукофитам ) . Упоминавшийся Кисом и Кремером род (также монотипический) в настоящее время относят не к глаукофитам, а к красным водорослям, где его выделяют в отдельный порядок класса . Впрочем, вероятно, видовое разнообразие глаукофитов недооценено: анализ генетических расхождений между отдельными представителями некоторых видов, выделяемых в настоящее время в родах Cyanophora и Glaucocystis , выявил столь значительные различия, что такие виды разумнее разделить на несколько самостоятельных видов ; в частности, на этом основании группа японских исследователей в 2014 году выделила 3 новых вида в роду Cyanophora .
Согласно наиболее распространённой точке зрения, отдел Глаукофитовые водоросли Glaucophyta (= Glaucocystophyta L.Kies & B.P.Kremer, 1986 ; по Т. Кавалье-Смиту — «ненужный синоним » ) включает один класс Glaucophyceae Bohlin, 1901 (= Glaucocystophyceae J.H.Schaffn., 1922 ) c единственным порядком Glaucocystales Bessey , 1907 и единственным же семейством Glaucocystaceae G.S.West , 1904 , содержащем все виды и роды глаукофитов .
Правда, Кис и Кремер выделяли в составе отдела 3 порядка (Cyanophorales, Glaucocystales, Gloeochaetales — соответственно для монадных, коккоидных и пальмеллоидных форм) и 4 семейства (иногда этой схеме следуют и современные источники ); однако эта классификация строилась на основе морфологических данных и к началу XXI века устарела. Построение надёжной современной классификации глаукофитовых водорослей, основанной на анализе их филогении и учитывающей не только морфологические, но и молекулярные данные , осложняется как противоречивостью полученных к настоящему времени результатов филогенетического анализа (так, в одном из исследований 2014 года роды Glaucocystis и Gloeochaete выступают — хотя и с невысокой надёжностью — как сестринские группы, но род Cyanophora в зависимости от выбранной методики предстаёт либо как наиболее рано отделившаяся ветвь глаукофитов, либо как сестринская группа для рода Cyanoptyche ), так и неполнотой данных: в 2014 году только 4 рода (уже упомянутые Cyanophora , Glaucocystis , Cyanoptyche , Gloeochaete ) были представлены в коллекциях культур и базах данных о геномах водорослей .
Ниже приведён — в соответствии с информацией из базы данных AlgaeBase ?! — перечень признаваемых в настоящее время родов и видов глаукофитов (в списках видов на первое место вынесен типовой вид ) :