Interested Article - Археи
- 2020-02-08
- 1
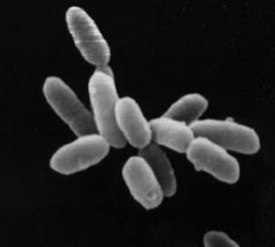
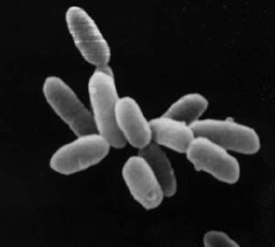
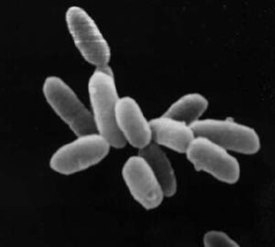
Архе́и (в единственном числе — архе́я , от лат. Archaea , от др.-греч. ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых организмов (по трёхдоменной системе Карла Вёзе наряду с бактериями и эукариотами ). Археи представляют собой одноклеточные микроорганизмы, не имеющие ядра , а также каких-либо мембранных органелл .
Ранее археи объединяли с бактериями в общую группу, называемую прокариоты (или царство Дробянки ( лат. Monera )), и они назывались архебактерии , однако сейчас такая классификация считается устаревшей : установлено, что археи имеют свою независимую эволюционную историю и характеризуются многими биохимическими особенностями, отличающими их от других форм жизни.
Сейчас археи подразделяют на более чем 7 типов . Из них наиболее изучены кренархеоты ( Crenarchaeota ) и эвриархеоты ( Euryarchaeota ). Классифицировать археи по-прежнему сложно, так как подавляющее большинство из них никогда не выращивались в лабораторных условиях и идентифицировались только по анализу нуклеиновых кислот из проб, полученных из мест их обитания.
Археи и бактерии очень похожи по размеру и форме клеток , хотя некоторые археи имеют довольно необычную форму, например, клетки Haloquadratum walsbyi плоские и квадратные. Несмотря на внешнее сходство с бактериями, некоторые гены и метаболические пути архей сближают их с эукариотами (в частности ферменты , катализирующие процессы транскрипции и трансляции ). Другие аспекты биохимии архей уникальны, к примеру, присутствие в клеточных мембранах липидов , содержащих простую эфирную связь . Большая часть архей — хемоавтотрофы . Они используют значительно больше источников энергии , чем эукариоты: начиная от обыкновенных органических соединений , таких как сахара , и заканчивая аммиаком , ионами металлов и даже водородом . Солеустойчивые археи — галоархеи ( Haloarchaea ) — используют в качестве источника энергии солнечный свет, другие виды архей фиксируют углерод , однако, в отличие от растений и цианобактерий (синезелёных водорослей), ни один вид архей не делает и то, и другое одновременно. Размножение у архей бесполое : бинарное деление , фрагментация и почкование . В отличие от бактерий и эукариот, ни один известный вид архей не формирует спор .
Изначально архей считали экстремофилами , живущими в суровых условиях — горячих источниках, солёных озёрах, однако потом их нашли и в более привычных местах, включая почву , океаны , болота и толстую кишку человека . Архей особенно много в океанах, и, возможно, планктонные археи — самая многочисленная группа ныне живущих организмов. Археи признаны важной составляющей жизни на Земле. Они играют роль в круговоротах углерода и азота . Ни один из известных представителей архей не является ни паразитом (за исключением наноархеот , являющихся паразитами других архей), ни патогенным организмом , однако они часто бывают мутуалистами и комменсалами . Некоторые представители являются метаногенами и обитают в пищеварительном тракте человека и жвачных , где они помогают осуществлять пищеварение. Метаногены используются в производстве биогаза и при очистке канализационных сточных вод, а ферменты экстремофильных микроорганизмов, сохраняющие активность при высоких температурах и в контакте с органическими растворителями, находят своё применение в биотехнологии .
История открытия

Первые представители группы обнаружены в различных экстремальных средах обитания , например, геотермальных источниках.
На протяжении большей части XX века прокариоты считались единой группой и классифицировались по биохимическим, морфологическим и метаболическим особенностям. К примеру, микробиологи пытались классифицировать микроорганизмы в зависимости от формы клеток, деталей строения клеточной стенки и потребляемых микроорганизмами веществ . В 1965 году было предложено устанавливать степень родства разных прокариот на основании сходства строения их генов . Этот подход, филогенетика , в наши дни является основным.
Впервые археи были выделены в качестве отдельной группы прокариот на филогенетическом древе в 1977 году Карлом Вёзе и Джорджем Эдвардом Фоксом при сравнительном анализе 16S рРНК . Изначально эти две группы обозначались как архебактерии ( лат. Archaebacteria ) и эубактерии ( лат. Eubacteria ) и рассматривались как царства или подцарства, которые Вёзе и Фокс называли термином Urkingdoms. Вёзе настаивал, что эта группа прокариот есть фундаментально отличный тип жизни. Чтобы подчеркнуть это отличие, впоследствии две группы прокариот были названы археями и бактериями . В трёхдоменной системе Карла Вёзе обе эти группы и эукариоты были возведены в ранг домена . Этот термин был предложен Вёзе в 1990 году для обозначения самого верхнего ранга в классификации организмов, включающей одно или несколько царств .
В первое время к новому домену причисляли только метаногенные микроорганизмы. Считалось, что археи населяют только места с экстремальными условиями: горячие источники , солёные озёра. Однако к концу XX века микробиологи пришли к выводу, что археи — большая и разнообразная группа организмов, широко распространённая в природе. Многие виды архей населяют вполне обычные среды обитания, например, почвы или воды океана . Такая переоценка была вызвана применением метода полимеразной цепной реакции для идентификации прокариот в образцах воды и почвы по их нуклеиновым кислотам . Данный метод позволяет выявлять и идентифицировать организмы, которые по тем или иным причинам не культивируются в лабораторных условиях .
Происхождение и эволюция
Хотя возможные окаменелости прокариотических клеток датированы возрастом в 3,5 млрд лет, большинство прокариот не имеет характерных морфологических особенностей, и поэтому окаменелые формы нельзя определённо идентифицировать именно как останки архей . В то же время химические остатки уникальных для архей липидов более информативны, так как эти соединения у других организмов не встречаются . В некоторых публикациях указывается, что останки липидов архей или эукариот присутствуют в породах возрастом 2,7 млрд лет , однако достоверность этих данных остаётся под сомнением . Эти липиды обнаружены в докембрийских формациях. Древнейшие из подобных остатков найдены в на западе Гренландии , где находятся самые старые на Земле осадочные породы , сформировавшиеся 3,8 млрд лет назад . Археи могут быть древнейшими живыми существами, населяющими Землю .
Вёзе утверждал, что археи, бактерии и эукариоты представляют собой три раздельные линии, рано отделившиеся от общей предковой группы организмов . Возможно, это произошло ещё до клеточной эволюции, когда отсутствие типичной клеточной мембраны давало возможности к неограниченному горизонтальному переносу генов , и предки трёх доменов различались между собой по фиксируемым комплектам генов . Не исключено, что последний общий предок архей и бактерий был термофилом , это даёт основания предположить, что низкие температуры были «экстремальной средой» для архей, и организмы, приспособившиеся к ним, появились только позже . Сейчас археи и бактерии связаны между собой не больше, чем с эукариотами, и термин «прокариоты» обозначает лишь «не эукариоты», что ограничивает его применимость .
Сравнительная характеристика архей и других доменов
В приведённой таблице показаны некоторые черты архей, свойственные и не свойственные другим доменам . Многие из этих свойств также обсуждаются ниже.
| Свойственно археям и бактериям | Свойственно археям и эукариотам | Свойственно только археям |
|---|---|---|
| Нет оформленного ядра и мембранных органелл | Нет пептидогликана (муреина) | Структура клеточной стенки (к примеру, клеточные стенки некоторых архей содержат ) |
| Кольцевая хромосома | ДНК связана с гистонами | В клеточной мембране присутствуют липиды, содержащие простую эфирную связь |
| Гены объединены в опероны | Трансляция начинается с метионина | Структура флагеллинов |
| Схожие РНК-полимераза , промоторы и другие компоненты транскрипционного комплекса, есть интроны и процессинг РНК | Структура рибосом (некоторые признаки сближают с бактериями, некоторые — с эукариотами) | |
| Полицистронная мРНК | Схожие репликация и репарация ДНК | Структура и метаболизм тРНК |
| Размер клеток на несколько порядков меньше, чем у эукариот | Схожая АТФаза (тип V) |
Родство с другими прокариотами
Установление степени родства между тремя доменами имеет ключевое значение для понимания возникновения жизни. Большинство метаболических путей, в которых задействована большая часть генов организма, схожи у бактерий и архей, в то время как гены, отвечающие за экспрессию других генов, очень похожи у архей и эукариот . По строению клеток археи наиболее близки к грамположительным бактериям : клетка покрыта единственной плазматической мембраной, дополнительная внешняя мембрана, характерная для грамотрицательных бактерий , отсутствует , клеточные стенки различного химического состава, как правило, толстые . В филогенетическом древе, основанном на сравнительном анализе структур гомологичных генов/белков прокариот, гомологи архей наиболее близки к таковым грамположительных бактерий . В некоторых важнейших белках архей и грамположительных бактерий, таких как Hsp70 и глутаминовая синтетаза I, обнаруживаются одинаковые эволюционно консервативные вставки и делеции .
Гупта ( англ. Gupta ) предположил, что археи отделились от грамположительных бактерий в результате отбора по признаку устойчивости к действию антибиотиков . Это основывается на наблюдении, что археи устойчивы к огромному числу антибиотиков , в основном производимых грамположительными бактериями , и что эти антибиотики действуют главным образом на гены, которые отличают бактерий от архей. Согласно гипотезе Гупты, давление отбора в направлении формирования устойчивости к антибиотикам грамположительных бактерий в конце концов привело к существенным изменениям в структуре генов-мишеней антибиотиков у некоторых микроорганизмов, которые стали общими предками современных архей . Предполагаемая эволюция архей под действием антибиотиков и других неблагоприятных факторов также может объяснить их адаптацию к экстремальным условиям, таким как повышенные температура и кислотность , как результат поиска ниш, свободных от продуцирующих антибиотики организмов . Т. Кавалир-Смит выдвинул схожее предположение . Версия Гупты также подтверждается другими работами, изучающими родственные черты в структурах белков , и исследованиями, показавшими, что грамположительные бактерии могли быть первой ветвью, отделившейся от общего древа прокариот .
Родство с эукариотами
Эволюционное родство между археями и эукариотами остаётся неясным. Помимо сходства в структуре и функциях клеток, между ними существует сходство на генетическом уровне. Установлено, что группа архей кренархеоты стоят ближе к эукариотам, чем к другому типу архей — эвриархеотам . Кроме того, у некоторых бактерий, как обнаружены архееподобные гены, переданные путём горизонтального переноса . Наиболее распространена гипотеза, согласно которой предок эукариот рано отделился от архей , а эукариоты возникли в результате слияния археи и эубактерии, ставших цитоплазмой и ядром новой клетки. Эта гипотеза объясняет различные генетические сходства, но сталкивается с трудностями в объяснении клеточной структуры .
Строение
Форма клеток и колоний
Отдельные клетки архей достигают от 0,1 до 15 мкм в диаметре и могут иметь различную форму: шара, палочки, спирали или диска . Некоторые кренархеоты имеют другую форму, например, — неправильной дольчатой формы; — тонкой нитевидной формы и меньше 1 мкм в диаметре, а и почти идеально прямоугольные . Haloquadratum walsbyi — плоские квадратные археи, живущие в сверхсолёных водоёмах . Такие необычные формы клеток, вероятно, обеспечиваются клеточной стенкой и прокариотическим цитоскелетом . У архей обнаружены белки, родственные компонентам цитоскелета других организмов , а также показано присутствие филаментов в их клетках , однако у архей, в отличие от других организмов, эти структуры плохо изучены . У Thermoplasma и Ferroplasma клеточная стенка отсутствует, поэтому их клетки имеют неправильную форму и похожи на амёб .
Клетки некоторых видов архей могут объединяться в агрегаты и филаменты длиной до 200 мкм. Эти организмы могут формировать биоплёнки . В культурах клетки сливаются друг с другом, формируя одну крупную клетку . Археи рода образуют сложные многоклеточные колонии, в которых клетки объединены с помощью длинных, тонких, полых трубок, называемых cannulae , которые выступают над поверхностями клеток и собирают их в густое кустовидное скопление . Функции этих трубок не ясны, но, возможно, они осуществляют коммуникацию и обмен питательными веществами между соседними клетками . Существуют и многовидовые колонии, как, например, «нить жемчуга», обнаруженная в 2001 году в болоте в Германии . Круглые беловатые колонии некоторых необычных эвриархеот перемежаются тонкими нитями, которые могут достигать до 15 см в длину и состоят из особых видов бактерий .
Археи и бактерии имеют очень похожую структуру клеток, однако их состав и организация отделяют архей от бактерий. Как у бактерий, у них отсутствуют внутренние мембраны и органеллы , клеточные мембраны, как правило, ограничены клеточной стенкой , а плавание осуществляется за счёт одного или более жгутиков . Структурно археи наиболее схожи с грамположительными бактериями. Большинство имеет одну плазматическую мембрану и клеточную стенку, периплазматическое пространство отсутствует. Исключением из этого главного правила является , у которого крупное периплазматическое пространство, ограниченное наружной мембраной, содержит окружённые мембраной везикулы .
Мембраны

Молекулы , из которых построены мембраны архей, сильно отличаются от тех, которые используются в мембранах других организмов. Это указывает на то, что археи состоят лишь в отдалённом родстве с бактериями и эукариотами . У всех живых организмов клеточные мембраны построены из фосфолипидов . Молекулы фосфолипидов состоят из двух частей: гидрофильной полярной , состоящей из фосфатов , и гидрофобной неполярной, состоящей из липидов . Эти компоненты объединены через остаток глицерина . В воде молекулы фосфолипидов кластеризуются, при этом фосфатные «головки» оказываются обращёнными к воде, а липидные «хвосты» — обращёнными от неё и спрятанными внутрь кластера. Главная составляющая мембраны — два слоя таких фосфолипидов, называемые липидным бислоем.
Эти фосфолипиды у архей обладают четырьмя необычными чертами:
- У бактерий и эукариот мембраны состоят главным образом из глицерин- сложноэфирных липидов, тогда как у архей они сложены из глицерин- эфирных липидов . Различается тип связи между остатками липидов и глицерина. Связи двух типов обозначены жёлтым на схеме справа. В сложноэфирных липидах связь сложноэфирная, а в эфирных — эфирная. Эфирные связи химически более стойкие, чем сложноэфирные. Эта стабильность помогает археям выживать при высоких температурах, а также в сильнокислых и сильнощелочных средах . Бактерии и эукариоты содержат некоторое количество эфирных липидов, но по сравнению с археями они не являются главной составляющей мембран.
- Имеется отличие в стереохимии — у архей асимметрический центр глицериновой составляющей имеет L- конфигурацию , а не D-, как у других организмов. Поэтому для синтеза фосфолипидов археи используют совершенно другие ферменты , чем бактерии и эукариоты. Такие ферменты появились очень рано в истории жизни, что указывает на то, что археи рано отделились от двух других доменов .
- Липидные «хвосты» архей химически отличны от таковых у других организмов. Основу липидов архей составляет изопреноидная боковая цепь, и их липиды представляют собой длинные цепи с множеством побочных ветвей, иногда даже с циклопропановыми и циклогексановыми кольцами . Хотя изопреноиды играют важную роль в биохимии многих организмов, только археи используют их для создания фосфолипидов. Предполагают, что эти разветвлённые цепи, как и эфирные связи, служат для приспособления к обитанию при высоких температурах. Установлено, что изопреноидные мембраны сохраняют в широком диапазоне температур (0—100 °C) жидкокристаллическое состояние, что необходимо для их нормального биологического функционирования. Проницаемость таких мембран для ионов и низкомолекулярных органических веществ также мало изменяется с повышением температуры, в отличие от мембран из «обычных» липидов, у которых она резко возрастает .
- У некоторых архей липидный бислой заменяется монослоем. Фактически при этом липидные «хвосты» двух разных фосфолипидных молекул сливаются с образованием одной молекулы с двумя полярными головками. Эти слияния делают мембрану более стойкой и лучше приспособленной для суровых условий . К примеру, ферроплазма имеет липиды этого типа, и они помогают ей выживать в сильнокислых условиях .
Клеточная стенка
Большинство архей (но не Thermoplasma и Ferroplasma ) обладают клеточной стенкой . У большей части из них она сформирована молекулами поверхностных белков, образующих наружный S-слой . S-слой представляет собой жёсткую сетку из белковых молекул, покрывающих клетку снаружи, подобно кольчуге . Этот слой защищает клетку от физических и химических воздействий, а также предотвращает контакт макромолекул с клеточной мембраной . В отличие от бактерий, клеточная стенка архей не содержит пептидогликан . ( лат. Methanobacteriales ) имеют клеточные стенки, содержащие , который напоминает пептидогликан эубактерий по морфологии, функции и физической структуре, но отличен по химической: в нём нет остатков D-аминокислот и N-ацетилмурамовой кислоты .
Жгутики
Жгутик архей иногда называют археллум . Жгутики архей работают так же, как и у бактерий: их длинные нити приводятся в движение вращательным механизмом в основании жгутика. Этот механизм работает за счёт трансмембранного протонного градиента. Тем не менее жгутики архей значительно отличаются от бактериальных по строению и способу сборки . Два типа жгутиков развились из разных предковых структур. Бактериальный жгутик и система секреции III типа имели общую предковую структуру , а архейный жгутик произошёл от бактериальных пилей IV типа . Жгутик бактерий полый и собирается из субъединиц, которые проходят вверх по центральной поре к концу жгутика. Жгутики же архей строятся путём добавления субъединиц в их основание . Кроме того, в отличие от бактериальных жгутиков, в жгутики архей входит несколько видов флагеллинов .
Метаболизм
Археи демонстрируют огромное разнообразие химических реакций, протекающих в их клетках в процессе метаболизма, а также источников энергии. Эти реакции классифицируются по группам питания в зависимости от источников энергии и углерода . Некоторые археи получают энергию из неорганических соединений , таких как сера или аммиак (они являются литотрофами ). К ним относятся нитрифицирующие археи, метаногены и анаэробные метаноокислители . В этих реакциях одно соединение отдаёт электроны другому ( окислительно-восстановительные реакции ), а выделяющаяся при этом энергия служит топливом для осуществления различных клеточных процессов. Соединение, отдающее электроны, называется донором, а принимающее — акцептором. Выделяющаяся энергия идёт на образование АТФ путём хемиосмоса . В сущности, это основной процесс, протекающий в митохондриях эукариотических клеток .
Другие группы архей используют в качестве источника энергии солнечный свет (их называют фототрофами ). Однако ни один из этих организмов не образует кислород в процессе фотосинтеза . Многие базовые метаболические процессы являются общими для всех форм жизни, например, археи используют модифицированный вариант гликолиза ( путь Энтнера-Дудорова ), а также полный или частичный цикл Кребса (трикарбоновых кислот) . Это, вероятно, отражает раннее возникновение этих путей в истории жизни и их высокую эффективность .
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические соединения | Halobacteria |
| Литотрофы | Неорганические соединения | Органические соединения или фиксация углерода | , Methanobacteria , |
| Органотрофы | Органические соединения | Органические соединения или фиксация углерода | , , Methanosarcinales |
Некоторые эвриархеоты являются метаногенами и обитают в анаэробных средах, таких как болота. Такой тип метаболизма появился рано, и возможно даже, что первый свободноживущий организм был метаногеном . Обычная для этих организмов биохимическая реакция представляет собой окисление водорода с использованием углекислого газа в качестве акцептора электронов. Для осуществления метаногенеза необходимо множество различных коферментов , уникальных для этих архей, таких как кофермент М и метанофуран . Некоторые органические соединения, такие как спирты , уксусная и муравьиная кислоты , могут использоваться метаногенами в качестве альтернативных акцепторов электронов. Подобные реакции протекают у архей, живущих в пищеварительном тракте . У ацидотрофных архей уксусная кислота распадается непосредственно на метан и углекислый газ. Такие ацидотрофные археи относятся к отряду Methanosarcinales . Они являются важной составляющей сообществ микроорганизмов, продуцирующих биогаз .
Другие археи используют атмосферный углекислый газ как источник углерода благодаря процессу фиксации углерода (то есть являются автотрофами ). Этот процесс включает в себя либо сильно изменённый цикл Кальвина , либо недавно открытый метаболический путь, известный как 3-гидроксилпропионат/4-гидроксибутиратный цикл . Кренархеоты также используют обратный цикл Кребса, а эвриархеоты — восстановительный ацетил-СоА процесс . Фиксация углерода осуществляется за счёт энергии, получаемой из неорганических соединений. Ни один известный вид архей не фотосинтезирует . Источники энергии, которые используют археи, чрезвычайно разнообразны, начиная от окисления аммиака до окисления сероводорода или элементарной серы, проводимого , при этом в качестве акцепторов электронов могут использоваться кислород или ионы металлов .

Фототрофные археи используют солнечный свет для получения химической энергии в виде АТФ . У Halobacteria активируемые светом ионные насосы как бактериородопсин и создают ионный градиент путём выкачивания ионов из клетки через плазматическую мембрану. Запасённая в этом электрохимическом градиенте энергия преобразуется в АТФ с помощью АТФ-синтазы . Этот процесс представляет собой форму фотофосфорилирования . Способность этих насосов переносить ионы через мембраны при освещении обусловлена изменениями, которые происходят в структуре ретинолового кофактора , скрытого в центре белка, под действием света .
Генетика
Как правило, археи имеют одиночную кольцевую хромосому , размер которой может достигать 5 751 492 пар нуклеотидов у Methanosarcina acetivorans , обладающей самым большим известным геномом среди архей. Одну десятую размера этого генома составляет геном с 490 885 парами нуклеотидов у Nanoarchaeum equitans , имеющего самый маленький известный геном среди архей; он содержит лишь 537 генов , кодирующих белки . Также у архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды . Возможно, плазмиды могут передаваться между клетками при физическом контакте, в ходе процесса сходного с конъюгацией бактерий .

Археи могут поражаться вирусами , содержащими двухцепочечную ДНК. Вирусы архей часто неродственны другим группам вирусов и имеют различные необычные формы, включая бутылки, крючки и капли . Эти вирусы были тщательно изучены на термофилах, в основном отрядов Sulfolobales и Thermoproteales . В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражающий галофильные археи . Защитные реакции архей против вирусов могут включать механизм, близкий к РНК-интерференции эукариот .
Археи генетически отличны от эукариот и бактерий, причём до 15 % белков, кодируемых одним геномом археи, уникальны для этого домена, хотя функции большинства этих белков неизвестны . Большая часть уникальных белков, функция которых известна, принадлежит эвриархеотам и задействована в метаногенезе. Белки, общие для архей, бактерий и эукариот, участвуют в основных клеточных функциях и касаются в основном транскрипции , трансляции и метаболизма нуклеотидов . К другим особенностям архей можно отнести организацию генов, выполняющих связанные функции (к примеру, гены, ответственные за разные этапы одного и того же метаболического процесса), в опероны и большие отличия в строении генов тРНК и их аминоацил-тРНК синтетаз .
Транскрипция и трансляция архей больше напоминают эти процессы в клетках эукариот, чем бактерий, причём РНК-полимераза и рибосомы архей очень близки к аналогичным структурам у эукариот . Хотя у архей есть лишь один тип РНК-полимеразы, по строению и функции в транскрипции она близка к РНК-полимеразе II эукариот, при этом схожие группы белков (главные факторы транскрипции) обеспечивают связывание РНК-полимеразы с промотором гена . В то же время другие факторы транскрипции архей более близки к таковым у бактерий . Процессинг РНК у архей проще, чем у эукариот, так как большинство генов архей не содержит интронов , хотя в генах их тРНК и рРНК их достаточно много , также они присутствуют в небольшом количестве генов, кодирующих белки .
Размножение
Археи размножаются бесполым путём : бинарным или множественным делением, фрагментацией или почкованием . Мейоза не происходит, поэтому даже если представители конкретного вида архей существуют более чем в одной форме, все они имеют одинаковый генетический материал . Клеточное деление определяется клеточным циклом : после того, как хромосома реплицировалась и две дочерние хромосомы разошлись, клетка делится . Детали изучены лишь у рода , но особенности его цикла очень схожи с таковыми и у эукариот, и у бактерий. Репликация хромосом начинается с множественных точек начала репликации с помощью ДНК-полимеразы , похожей на аналогичные ферменты эукариот . Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки, и компоненты септы, проходящей через центр клетки, схожи с их бактериальными эквивалентами .
Археи не образуют споры . Некоторые виды могут претерпевать смену фенотипа и существовать как клетки нескольких различных типов, включая толстостенные клетки, устойчивые к осмотическому шоку и позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания .
Экология

Среды обитания
Археи живут в широком диапазоне сред обитания и являются важной частью глобальной экосистемы , могут составлять до 20 % общей биомассы . Первые открытые археи были экстремофилами . Действительно, многие археи выживают при высоких температурах , часто свыше 100 °C, и обнаружены в гейзерах , чёрных курильщиках и маслосборниках. Другие приспособились к жизни в очень холодных условиях, в сильносолёных, сильнокислых и сильнощелочных средах, а также при высоком давлении — до 700 атмосфер ( барофилы ). Однако среди архей есть и мезофилы , обитающие в мягких условиях, в болотистых местностях, сточных водах, океанах и почве .
Экстремофильные археи относятся к четырём главным физиологическим группам: галофилам , термофилам , ацидофилам (кислотоустойчивые) и алкалифилам (щелочеустойчивые) . Эти группы нельзя рассматривать в ранге типа или как другие самостоятельные таксоны. Они не взаимоисключают друг друга, и некоторые археи относят одновременно к нескольким группам. Тем не менее, они являются удобной стартовой точкой для классификации.
Галофилы, включая род , живут в экстремально солёных средах, таких как солёные озёра, и при минерализации больше 20—25 % превосходят по численности своих соседей-бактерий . Термофилы лучше всего растут на температурах свыше 45 °C в таких местах, как горячие источники; для гипертермофилов оптимальная температура — 80 °C и выше . Methanopyrus kandleri ( штамм 116) растёт при 122 °C, рекордно высокой температуре для всех организмов .
Другие археи обитают в очень кислых или щелочных средах . Например, наиболее устойчивый ацидофил растёт при pH = 0, что эквивалентно 1,2 молярной серной кислоте .
Устойчивость к экстремальным условиям внешней среды сделала архей центральной темой в обсуждениях возможных свойств жизни на других планетах . Некоторые среды, в которых обитают экстремофилы, не сильно отличаются от таковых на Марсе , что наводит на мысль о возможном переносе таких устойчивых микроорганизмов между планетами на метеоритах .
Недавно несколько работ показали, что археи обитают не только в термофильных и мезофильных условиях, но также встречаются, иногда в большом количестве, и в местах с низкими температурами. Например, археи встречаются в холодных водах, таких как полярные моря . Ещё более важно, что огромное количество архей обнаружено повсеместно в океанах в неэкстремальных условиях в составе планктона (как часть пикопланктона) . Хотя эти археи могут присутствовать в поистине колоссальном количестве (до 40 % от общей биомассы микробов), почти ни один из этих видов не был изолирован, выращен и изучен в чистой культуре . Поэтому наше понимание роли архей в экологии океана, их влияния на глобальный биогеохимический круговорот остаётся в значительной мере неполным . Некоторые морские кренархеоты способны к нитрификации , поэтому вероятно, что они оказывают влияние на океанический круговорот азота , хотя эти океанические кренархеоты могут использовать и другие источники энергии . Большое число архей также обнаружено в осадке, покрывающем океаническое дно, причём они составляют большинство живых клеток на глубине больше 1 м от уровня океанического дна .
Роль в круговороте веществ
Археи вторично используют такие элементы, как углерод , азот и серу в своих различных средах обитания. Хотя такие превращения необходимы для нормального функционирования экосистемы , археи могут также содействовать вредным изменениям, вызванным деятельностью человека, и даже вызвать загрязнение.
Археи осуществляют многие этапы круговорота азота . Это включает в себя как реакции, удаляющие азот из экосистемы, к примеру, азотное дыхание и денитрификация , так и процессы, в ходе которых поглощается азот, такие как усвоение нитратов и фиксация азота . Недавно была открыта причастность архей к окислению аммиака . Эти реакции особенно важны в океанах . Археи также играют важную роль в почвенном окислении аммиака. Они образуют нитриты , которые затем окисляются другими микробами в нитраты . Последние потребляются растениями и другими организмами .
В круговороте серы археи, живущие за счёт окисления соединений серы, получают их из каменистых пород и делают их доступными для других организмов. Однако виды, осуществляющие это, такие как , образуют серную кислоту как побочный продукт, и существование таких организмов в заброшенных шахтах может, совместно с кислотными шахтными водами, причинить вред окружающей среде .
В круговороте углерода метаногены удаляют водород и играют важную роль в разложении органической материи популяциями микроорганизмов, выступающих как разлагатели в анаэробных экосистемах, таких как илы, болота и водоочистные сооружения . Однако метан — один из самых распространённых газов в земной атмосфере , вызывающих парниковый эффект , достигая 18 % от общего объёма парниковых газов . Он в 25 раз более эффективен по способности вызывать парниковый эффект, чем углекислый газ . Метаногены (главный источник атмосферного метана) выделяют большую часть ежегодного выброса метана . Поэтому эти археи причастны к созданию парникового эффекта на Земле и глобальному потеплению .
Взаимодействие с другими организмами

Хорошо изученные отношения между археями и другими организмами — мутуализм и комменсализм . Пока не существует чётких доказательств существования патогенных или паразитических видов архей . Однако была предположена связь между некоторыми видами метаногенов и инфекциями полости рта . Кроме того, вид Nanoarchaeum equitans , возможно, является паразитом другого вида архей, поскольку он выживает и размножается только на клетках кренархеота и не приносит никакой очевидной выгоды своему хозяину . С другой стороны, архееподобные ацидофильные наноорганизмы Ричмондских рудников ( ARMAN ) иногда прикрепляются к клеткам других архей в биоплёнках кислых сточных вод рудников . Природа этого взаимодействия не ясна, но, в отличие от случая Nanorchaeaum—Ignicoccus, сверхмелкие клетки ARMAN всегда остаются независимыми от клеток Thermoplasmatales .
Мутуализм
Один из хорошо понятных примеров мутуализма — взаимодействие простейших и метанообразующих архей, обитающих в пищеварительном тракте животных , способных переваривать целлюлозу , таких как жвачные и термиты . В этих анаэробных условиях простейшие разлагают целлюлозу для получения энергии. В этом процессе в качестве побочного продукта освобождается водород , однако высокий его уровень сокращает получение энергии. Метаногены превращают водород в метан , и простейшие могут дальше нормально получать энергию .
В случае анаэробных простейших вроде археи живут внутри клетки простейшего и потребляют водород, образуемый в его гидрогеносомах . Археи также взаимодействуют и с более крупными организмами. Например, морская архея живёт внутри (как эндосимбионт) губки .
Комменсализм
Археи могут быть комменсалами , то есть существовать совместно с другим организмом, не принося ему ни пользы, ни вреда, но с выгодой для себя. К примеру, метаноген — наиболее типичный представитель архей в микрофлоре человека . Каждый десятый прокариот в человеческом пищеварительном тракте принадлежит к этому виду . В пищеварительном тракте термитов и человека эти метаногены в действительности могут быть мутуалистами , взаимодействующими с другими микробами пищеварительного тракта и способствующими пищеварению . Археи также взаимодействуют с другими организмами, к примеру, живут на внешней поверхности кораллов и в части почвы, прилегающей к корням растений ( ризосфере ) .
Классификация

Классификация архей, как и прокариот в целом, быстро меняется и во многом остаётся спорной. Современные системы классификации стремятся объединить археи в группы организмов со схожими структурными свойствами и общими предками . Эти классификации основаны на анализе структуры генов рРНК для установления родственных отношений между организмами ( молекулярная филогенетика ) . Большую часть архей, выращиваемых в лабораториях и хорошо изученных, относят к двум главным типам, кренархеоты ( Crenarchaeota ) и эвриархеоты ( Euryarchaeota ). Другие группы были выделены в порядке рабочей гипотезы. Например, довольно необычный вид Nanoarchaeum equitans , открытый в 2003 году, был выделен в самостоятельный тип Nanoarchaeota . Был также предложен новый тип Korarchaeota . Он объединяет небольшую группу термофильных видов, обладающих особенностями обоих основных типов, но более родственно близких к кренархеотам . Другие недавно открытые виды имеют лишь дальнее родство с вышеперечисленными группами, например, архейные ацидофильные наноорганизмы Ричмондских рудников (ARMAN) , открытые в 2006 году и являющиеся одними из самых мелких известных на сегодняшний момент организмов .

Разделение архей на виды также спорно. В биологии вид определяется как группа близкородственных организмов. Обычный критерий, которым пользуются в подобных ситуациях — организмы одного вида могут скрещиваться друг с другом, но не с особями других видов, — в данном случае не работает, поскольку археи размножаются только бесполым путём .
Археи демонстрируют высокий уровень горизонтального переноса генов между линиями. Некоторые исследователи предполагают, что особи можно объединять в популяции, похожие на виды, при условии высокой степени схожести их геномов и редко случающегося переноса генов между организмами с менее схожими геномами, как в случае рода ферроплазма ( Ferroplasma ) . С другой стороны, изучение рода показало существование значимой передачи генов между дальнеродственными популяциями, что ограничивает применимость этого критерия . Вторая проблема состоит в том, какое практическое значение может иметь подобное разделение на виды .
Современные данные о генетическом разнообразии архей фрагментарны, и общее число их видов не может быть оценено с какой-либо точностью . Сравнительный анализ структур 16S рРНК архей позволил предположить существование 18—23 филогенетических групп уровня типов , причём представители лишь восьми групп выращены непосредственно в лаборатории и изучены (с учётом вероятной полифилии некоторых выделяемых в данный момент типов). Многие из этих гипотетических групп известны лишь по одной последовательности рРНК, что говорит о том, что пределы разнообразия этих организмов остаются неясными . Многие бактерии также никогда не культивировались в лаборатории, что приводит к схожим проблемам при их характеристике .
По устоявшейся классификации на апрель 2021 года выделяют не менее 12 типов архей :
- Crenarchaeota Garrity and Holt 2001 — Кренархеоты — термофилы, термоацидофилы, серные анаэробные бактерии ;
- Euryarchaeota Garrity and Holt 2001 — Эвриархеоты — метаногенные и галофильные археи;
- Thaumarchaeota Brochier-Armanet et al. 2008 — в основном окислители аммония, как например, морской аммоний-окислитель и аммоний-окислитель преимущественно почвенного происхождения . Недавние филогенетические исследования, основанные на сравнительном анализе структур рибосомальных белков и других важнейших генов, подтвердили существование этого типа ;
- Candidatus Aigarchaeota Nunoura et al. 2011
- Candidatus Rinke et al. 2013
- Candidatus Korarchaeota Barns et al. 1996 — Корархеоты — ДНК обнаружена в геотермальных источниках США , Исландии , на рисовых полях Японии , культивируемые виды пока неизвестны;
- Candidatus Lokiarchaeota Spang et al. 2015 — наиболее известный представитель, , выделен на основании генома, собранного при метагеномном анализе образцов, полученных рядом с гидротермальными источниками в Атлантическом океане на глубине 2,35 км ;
- Candidatus Nanoarchaeota Huber et al. 2002 — Наноархеоты — единственные известные представители Nanoarchaeum equitans и ;
- Candidatus Rinke et al. 2013
- Candidatus Rinke et al. 2013
- Candidatus Dombrowski et al. 2020
- Candidatus Vanwonterghem et al. 2016
Выделяют ещё несколько типов на основании данных метагеномики , в том числе:

Филогенетический анализ показал, что Lokiarchaeota и эукариоты образуют монофилетическую кладу — в их геномах обнаружены близкие гены, например, гены, кодирующие белки, отвечающие за изменение формы клеточной мембраны, определение формы клетки и динамический цитоскелет . Результаты этого исследования служат подтверждением так называемой двухдоменной, или , согласно которой эукариоты появились как особая группа внутри архей, близкая к Lokiarchaeota и приобретшая митохондрии в результате эндосимбиоза .
В январе 2016 года были опубликованы результаты метагеномных исследований по реконструкции геномов архей из морских осадочных отложений, которые свидетельствуют об обнаружении нового типа архей — Thorarchaeota . Организмы этой группы способны к образованию ацетата при деградации белков. Они также имеют гены, необходимые для восстановления элементарной серы и тиосульфата , поэтому эти организмы участвуют в круговороте серы .
Значение в технологии и промышленности
Экстремофильные археи, особенно устойчивые к высоким температурам или повышенной кислотности/щёлочности среды, являются источником ферментов , работающих в этих суровых условиях . Эти ферменты находят множество применений. Например, термостабильные ДНК-полимеразы , такие как Pfu ДНК-полимераза вида , полностью изменили молекулярную биологию, дав возможность использовать полимеразную цепную реакцию для простого и быстрого клонирования ДНК . В промышленности амилазы , и других видов , функционирующие при температуре свыше 100 °C, применяются при производстве продуктов питания при высоких температурах, к примеру, при производстве молока и сыворотки с низким содержанием лактозы . Ферменты этих термофильных архей остаются очень стабильными в органических растворителях , что позволяет использовать их в безопасных для окружающей среды процессах в зелёной химии для синтеза органических соединений . Их стабильность делает эти ферменты удобными для использования в структурной биологии , поэтому аналоги ферментов бактерий и эукариот, получаемые из экстремофильных архей, часто применяются в структурных исследованиях .
По сравнению с применением ферментов архей, использование самих организмов в биотехнологии развито слабо. Метанообразующие археи — важная часть водоочистных сооружений, так как они входят в сообщество микроорганизмов, осуществляющих анаэробное разложение и образование биогаза . В обогащении полезных ископаемых ацидофильные археи могут использоваться для получения металлов из руд , в том числе золота , кобальта и меди .
Археи могут дать потенциально полезные антибиотики . Пока описано мало , но предполагается, что их существует сотни, особенно много их может быть получено из родов и . Эти антибиотики по структуре отличны от бактериальных, поэтому они могут иметь другие механизмы действия. Кроме того, они могут позволить создать селектируемые маркеры для использования в молекулярной биологии архей .
Примечания
- (англ.) на сайте Национального центра биотехнологической информации (NCBI). (Дата обращения: 3 сентября 2020) .
- ↑ : [ англ. ] // . — . (Дата обращения: 16 мая 2021) .
- / Бонч-Осмоловская Е. А. // Анкилоз — Банка. — М. : Большая российская энциклопедия, 2005. — С. 311—312. — ( Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 2). — ISBN 5-85270-330-3 .
- Pace N. R. Time for a change (англ.) // Nature. — 2006. — May ( vol. 441 , no. 7091 ). — P. 289 . — doi : . — . — .
- . Дата обращения: 25 июля 2012. 30 августа 2014 года.
- Staley J. T. (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361 , no. 1475 . — P. 1899—1909 . — doi : . — . — PMC .
- Zuckerkandl E., Pauling L. Molecules as documents of evolutionary history (англ.) // : journal. — 1965. — Vol. 8 , no. 2 . — P. 357—366 . — doi : . — .
- Woese C. R., Kandler O., Wheelis M. L. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1990. — Vol. 87 , no. 12 . — P. 4576—4579 . — doi : . — . — . — PMC . 16 октября 2019 года.
- Woese C.R., Kandler O., Wheelis M.L. // Proc. Natl. Acad. Sci. USA. — 1990. — Т. 87 . — С. 4576—4579 . 9 июля 2008 года.
- ↑ DeLong E. F. Everything in moderation: archaea as 'non-extremophiles' (англ.) // Curr. Opin. Genet. Dev. : journal. — 1998. — Vol. 8 , no. 6 . — P. 649—654 . — doi : . — .
- Theron J., Cloete T. E. Molecular techniques for determining microbial diversity and community structure in natural environments (англ.) // : journal. — 2000. — Vol. 26 , no. 1 . — P. 37—57 . — doi : . — .
- Schmidt T. M. (англ.) // : journal. — 2006. — Vol. 9 , no. 3 . — P. 217—223 . — . 11 сентября 2008 года.
- Schopf J. (англ.) // Philos Trans R Soc Lond B Biol Sci : journal. — 2006. — Vol. 361 , no. 1470 . — P. 869—885 . — doi : . — . — PMC . 23 февраля 2011 года.
- Chappe B., Albrecht P., Michaelis W. Polar Lipids of Archaebacteria in Sediments and Petroleums (англ.) // Science : journal. — 1982. — July ( vol. 217 , no. 4554 ). — P. 65—66 . — doi : . — . — .
- Brocks J. J., Logan G. A., Buick R., Summons R. E. Archean molecular fossils and the early rise of eukaryotes (англ.) // Science : journal. — 1999. — Vol. 285 , no. 5430 . — P. 1033—1036 . — doi : . — .
- Rasmussen B., Fletcher I. R., Brocks J. J., Kilburn M. R. Reassessing the first appearance of eukaryotes and cyanobacteria (англ.) // Nature : journal. — 2008. — October ( vol. 455 , no. 7216 ). — P. 1101—1104 . — doi : . — . — .
- Hahn, Jürgen; Pat Haug. Traces of Archaebacteria in ancient sediments (англ.) // System Applied Microbiology. — 1986. — Vol. 7 , no. Archaebacteria '85 Proceedings . — P. 178—183 .
- Wang M., Yafremava L. S., Caetano-Anollés D., Mittenthal J. E., Caetano-Anollés G. Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world (англ.) // : journal. — 2007. — Vol. 17 , no. 11 . — P. 1572—1585 . — doi : . — . — PMC .
- Woese C. R., Gupta R. Are archaebacteria merely derived 'prokaryotes'? (англ.) // Nature. — 1981. — Vol. 289 , no. 5793 . — P. 95—6 . — doi : . — . — .
- ↑ Woese C. R. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1998. — Vol. 95 , no. 12 . — P. 6854—6859 . — doi : . — . — . — PMC . 18 сентября 2019 года.
- Kandler O. The early diversification of life and the origin of the three domains: A proposal. In: Wiegel J., Adams W.W., editors. Thermophiles: The keys to molecular evolution and the origin of life? Athens: Taylor and Francis, 1998: 19-31.
- Gribaldo S., Brochier-Armanet C. (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361 , no. 1470 . — P. 1007—1022 . — doi : . — . — PMC . 4 июня 2012 года. . Дата обращения: 21 июля 2012. Архивировано из 4 июня 2012 года.
- ↑ Woese C. R. (англ.) // : journal. — , 1994. — 1 March ( vol. 58 , no. 1 ). — P. 1—9 . — . — PMC . 18 сентября 2019 года.
- Information is from Willey J.M., Sherwood L.M., Woolverton C.J. Microbiology 7th ed. (2008), Ch. 19 pp. 474—475, except where noted.
- Talbert P. B., Henikoff S. Histone variants – ancient wrap artists of the epigenome (англ.) // Nature Reviews Molecular Cell Biology : journal. — 2010. — Vol. 11 . — P. 264—275 . — doi : .
- Sandman K., Reeve J. N. Archaeal histones and the origin of the histone fold (англ.) // Curr. Opin. Microbiol : journal. — 2006. — Vol. 9 . — P. 520—525 . — doi : .
- у бактерий трансляция начинается с формилметионина
- ↑ Zillig W. Comparative biochemistry of Archaea and Bacteria (англ.) // Curr. Opin. Gen. Dev.. — 1991. — December ( vol. 1 , no. 4 ). — P. 544—551 . — doi : . — .
- Bell S. D., Jackson S. P. Mechanism and regulation of transcription in archaea (англ.) // Curr. Opin. Microbiol. : journal. — 2001. — April ( vol. 4 , no. 2 ). — P. 208—213 . — doi : . — .
- Reeve J. N. Archaeal chromatin and transcription (англ.) // : journal. — , 2003. — May ( vol. 48 , no. 3 ). — P. 587—598 . — .
- Kelman L. M., Kelman Z. Archaea: an archetype for replication initiation studies? (англ.) // : journal. — , 2003. — May ( vol. 48 , no. 3 ). — P. 605—615 . — .
- Phillips G., Chikwana V. M., Maxwell A., et al. Discovery and characterization of an amidinotransferase involved in the modification of archaeal tRNA (англ.) // J. Biol. Chem. : journal. — 2010. — April ( vol. 285 , no. 17 ). — P. 12706—12713 . — doi : . — . — PMC .
- Koonin E.V., Mushegian A.R., Galperin M.Y., Walker D.R. Comparison of archaeal and bacterial genomes: computer analysis of protein sequences predicts novel functions and suggests a chimeric origin for the archaea. Mol Microbiol 1997; 25:619-637.
- ↑ Gupta R. S. (англ.) // : journal. — , 1998. — Vol. 62 . — P. 1435—1491 . 15 марта 2021 года.
- Koch A.L. Were Gram-positive rods the first bacteria? Trends Microbiol 2003; 11(4):166-170.
- ↑ Gupta R. S. (англ.) // Mol. Microbiol : journal. — 1998. — Vol. 29 . — P. 695—708 . 6 июля 2017 года.
- Brown J.R., Masuchi Y., Robb F.T., Doolittle W.F. Evolutionary relationships of bacterial and archaeal glutamine synthetase genes. J Mol Evol 1994; 38(6):566-576.
- ↑ Gupta, R.S.(2000) The natural evolutionary relationships among prokaryotes. Crit. Rev. Microbiol. 26: 111—131.
- Gupta R.S. Molecular Sequences and the Early History of Life. In: Sapp J., editor. Microbial Phylogeny and Evolution: Concepts and Controversies. New York: Oxford University Press, 2005: 160—183.
- Cavalier-Smith T. The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. Int J Syst Evol Microbiol 2002; 52(Pt 1):7-76.
- Valas R.E., Bourne P.E.: 2011 The origin of a derived superkingdom: how a Gram-positive bacterium crossed the desert to become an archaeon. Biol Direct 6: 16.
- Skophammer R.G., Herbold C.W., Rivera M.C., Servin J.A., Lake J.A. Evidence that the root of the tree of life is not within the Archaea. Mol Biol Evol 2006; 23(9):1648-1651.
- Lake J. A. Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences (англ.) // Nature : journal. — 1988. — January ( vol. 331 , no. 6152 ). — P. 184—186 . — doi : . — . — .
- Nelson K. E., Clayton R. A., Gill S. R., ; Utterback, T.R.; Malek, J.A.; Linher, K.D.; Garrett, M.M.; Stewart, A.M.; Cotton, M.D.; Pratt, M.S.; Phillips, C.A.; Richardson, D.; Heidelberg, J.; Sutton, G.G.; Fleischmann, R.D.; Eisen, J.A.; White, O.; Salzberg, S.L.; Smith, H.O.; Venter, J.C.; Fraser, C.M. et al. Evidence for lateral gene transfer between Archaea and bacteria from genome sequence of Thermotoga maritima (англ.) // Nature : journal. — 1999. — Vol. 399 , no. 6734 . — P. 323—329 . — doi : . — . — .
- Gouy M., Li W. H. Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree (англ.) // Nature : journal. — 1989. — May ( vol. 339 , no. 6220 ). — P. 145—147 . — doi : . — . — .
- Yutin N., Makarova K. S., Mekhedov S. L., Wolf Y.I., Koonin E.V. (англ.) // : journal. — Oxford University Press , 2008. — May ( vol. 25 , no. 8 ). — P. 1619—1630 . — doi : . — . — PMC . 3 мая 2009 года.
- Lake J. A. Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences (англ.) // Nature : journal. — 1988. — Vol. 331 , no. 6152 . — P. 184—186 . — doi : . — . — .
- ↑ Krieg, Noel. (англ.) . — US: Springer, 2005. — P. —6. — ISBN 978-0-387-24143-2 .
- Barns, Sue and Burggraf, Siegfried. (1997) от 2 мая 2012 на Wayback Machine . Version 01 January 1997. in The Tree of Life Web Project
- Walsby, A. E. A square bacterium (англ.) // Nature. — 1980. — Vol. 283 , no. 5742 . — P. 69—71 . — doi : . — .
- Hara F., Yamashiro K., Nemoto N., et al. (англ.) // : journal. — 2007. — Vol. 189 , no. 5 . — P. 2039—2045 . — doi : . — . — PMC . 27 мая 2020 года.
- Trent J. D., Kagawa H. K., Yaoi T., Olle E., Zaluzec N. J. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1997. — Vol. 94 , no. 10 . — P. 5383—5388 . — doi : . — . — . — PMC . 27 мая 2020 года.
- Hixon W. G., Searcy D. G. Cytoskeleton in the archaebacterium Thermoplasma acidophilum? Viscosity increase in soluble extracts (англ.) // BioSystems : journal. — 1993. — Vol. 29 , no. 2—3 . — P. 151—160 . — doi : . — .
- ↑ Golyshina O. V., Pivovarova T. A., Karavaiko G. I., et al. (англ.) // : journal. — 2000. — 1 May ( vol. 50 , no. 3 ). — P. 997—1006 . — . (недоступная ссылка)
- Hall-Stoodley L., Costerton J. W., Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases (англ.) // Nat. Rev. Microbiol. : journal. — 2004. — Vol. 2 , no. 2 . — P. 95—108 . — doi : . — .
- Kuwabara T., Minaba M., Iwayama Y., ; Kamekura, M. et al. (англ.) // : journal. — 2005. — November ( vol. 55 , no. Pt 6 ). — P. 2507—2514 . — doi : . — . (недоступная ссылка)
- Nickell S., Hegerl R., Baumeister W., Rachel R. (англ.) // : journal. — 2003. — Vol. 141 , no. 1 . — P. 34—42 . — doi : . — . 27 июня 2018 года.
- Horn C., Paulmann B., Kerlen G., Junker N., Huber H. (англ.) // : journal. — 1999. — 15 August ( vol. 181 , no. 16 ). — P. 5114—5118 . — . — PMC . 16 октября 2019 года.
- Rudolph C., Wanner G., Huber R. Natural communities of novel archaea and bacteria growing in cold sulfurous springs with a string-of-pearls-like morphology (англ.) // : journal. — 2001. — May ( vol. 67 , no. 5 ). — P. 2336—2344 . — doi : . — . — PMC .
- ↑ Thomas N. A., Bardy S. L., Jarrell K. F. The archaeal flagellum: a different kind of prokaryotic motility structure (англ.) // : journal. — , 2001. — Vol. 25 , no. 2 . — P. 147—174 . — doi : . — .
- Rachel R., Wyschkony I., Riehl S., Huber H. (англ.) // Archaea : journal. — 2002. — March ( vol. 1 , no. 1 ). — P. 9—18 . — doi : . — . — PMC . 24 февраля 2009 года. . Дата обращения: 21 июля 2012. Архивировано из 24 февраля 2009 года.
- ↑ Koga Y., Morii H. (англ.) // : journal. — , 2007. — Vol. 71 , no. 1 . — P. 97—120 . — doi : . — . — PMC . 27 мая 2020 года.
- De Rosa M., Gambacorta A., Gliozzi A. (англ.) // : journal. — , 1986. — 1 March ( vol. 50 , no. 1 ). — P. 70—80 . — . — PMC . 16 октября 2019 года.
- Albers S. V., van de Vossenberg J. L., Driessen A. J., Konings W. N. (англ.) // : journal. — , 2000. — September ( vol. 5 ). — P. D813—20 . — doi : . — . 27 октября 2012 года.
- Damsté J. S., Schouten S., Hopmans E. C., van Duin A. C., Geenevasen J. A. (англ.) // : journal. — 2002. — October ( vol. 43 , no. 10 ). — P. 1641—1651 . — doi : . — . 27 мая 2020 года.
- Koga Y., Morii H. (англ.) // : journal. — 2005. — November ( vol. 69 , no. 11 ). — P. 2019—2034 . — doi : . — . 31 декабря 2008 года. . Дата обращения: 21 июля 2012. Архивировано 31 декабря 2008 года.
- Hanford M. J., Peeples T. L. Archaeal tetraether lipids: unique structures and applications (англ.) // Appl. Biochem. Biotechnol.. — 2002. — January ( vol. 97 , no. 1 ). — P. 45—62 . — doi : . — .
- Macalady J. L., Vestling M. M., Baumler D., Boekelheide N., Kaspar C. W., Banfield J. F. Tetraether-linked membrane monolayers in Ferroplasma spp: a key to survival in acid (англ.) // Extremophiles : journal. — 2004. — October ( vol. 8 , no. 5 ). — P. 411—419 . — doi : . — .
- Sára M., Sleytr U. B. (англ.) // : journal. — 2000. — Vol. 182 , no. 4 . — P. 859—868 . — doi : . — . — PMC . 27 мая 2020 года.
- Engelhardt H., Peters J. Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions (англ.) // : journal. — 1998. — Vol. 124 , no. 2—3 . — P. 276—302 . — doi : . — .
- ↑ Kandler, O.; König, H. (англ.) // Cellular and Molecular Life Sciences (CMLS). — 1998. — Vol. 54 , no. 4 . — P. 305—308 . — doi : . (недоступная ссылка)
- Howland, John L. (англ.) . — Oxford: Oxford University Press , 2000. — P. . — ISBN 0-19-511183-4 .
- Albers Sonja-Verena , Jarrell Ken F. (англ.) // Frontiers in Microbiology. — 2015. — 27 January ( vol. 6 ). — ISSN . — doi : .
- Gophna U., Ron E. Z., Graur D. (англ.) // : journal. — Elsevier , 2003. — July ( vol. 312 ). — P. 151—163 . — doi : . — . 22 ноября 2017 года.
- Nguyen L., Paulsen I. T., Tchieu J., Hueck C. J., Saier M. H. Phylogenetic analyses of the constituents of Type III protein secretion systems (англ.) // J. Mol. Microbiol. Biotechnol. : journal. — 2000. — April ( vol. 2 , no. 2 ). — P. 125—144 . — .
- Ng S. Y., Chaban B., Jarrell K. F. Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications (англ.) // J. Mol. Microbiol. Biotechnol. : journal. — 2006. — Vol. 11 , no. 3—5 . — P. 167—191 . — doi : . — .
- Bardy S. L., Ng S. Y., Jarrell K. F. (англ.) // : journal. — , 2003. — February ( vol. 149 , no. Pt 2 ). — P. 295—304 . — doi : . — .
- ↑ Valentine D. L. Adaptations to energy stress dictate the ecology and evolution of the Archaea (англ.) // Nat. Rev. Microbiol. : journal. — 2007. — Vol. 5 , no. 4 . — P. 316—323 . — doi : . — .
- ↑ Schäfer G., Engelhard M., Müller V. (англ.) // : journal. — , 1999. — 1 September ( vol. 63 , no. 3 ). — P. 570—620 . — . — PMC . 16 октября 2019 года.
- Romano A., Conway T. Evolution of carbohydrate metabolic pathways (англ.) // Res Microbiol. — 1996. — Vol. 147 , no. 6—7 . — P. 448—455 . — doi : . — .
- Koch A. How did bacteria come to be? (англ.) // : journal. — Academic Press , 1998. — Vol. 40 . — P. 353—399 . — doi : . — .
- DiMarco A. A., Bobik T. A., Wolfe R. S. Unusual coenzymes of methanogenesis (англ.) // : journal. — 1990. — Vol. 59 . — P. 355—394 . — doi : . — .
- Klocke M., Nettmann E., Bergmann I., et al. Characterization of the methanogenic Archaea within two-phase biogas reactor systems operated with plant biomass (англ.) // : journal. — 2008. — May ( vol. 31 , no. 3 ). — P. 190—205 . — doi : . — .
- Mueller-Cajar O., Badger M. R. New roads lead to Rubisco in archaebacteria (англ.) // : journal. — 2007. — August ( vol. 29 , no. 8 ). — P. 722—724 . — doi : . — .
- Berg I. A., Kockelkorn D., Buckel W., Fuchs G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea (англ.) // Science : journal. — 2007. — December ( vol. 318 , no. 5857 ). — P. 1782—1786 . — doi : . — . — .
- Thauer R. K. Microbiology. A fifth pathway of carbon fixation (англ.) // Science. — 2007. — December ( vol. 318 , no. 5857 ). — P. 1732—1733 . — doi : . — .
- Bryant D. A., Frigaard N. U. Prokaryotic photosynthesis and phototrophy illuminated (англ.) // : journal. — 2006. — November ( vol. 14 , no. 11 ). — P. 488—496 . — doi : . — .
- Könneke M., Bernhard A. E., de la Torre J. R., Walker C. B., Waterbury J. B., Stahl D. A. Isolation of an autotrophic ammonia-oxidizing marine archaeon (англ.) // Nature : journal. — 2005. — September ( vol. 437 , no. 7058 ). — P. 543—546 . — doi : . — . — .
- ↑ Francis C. A., Beman J. M., Kuypers M. M. New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation (англ.) // : journal. — 2007. — May ( vol. 1 , no. 1 ). — P. 19—27 . — doi : . — .
- Основано на от 3 марта 2016 на Wayback Machine . Данные опубликованы в Subramaniam S., Henderson R. Molecular mechanism of vectorial proton translocation by bacteriorhodopsin (англ.) // Nature : journal. — 2000. — August ( vol. 406 , no. 6796 ). — P. 653—657 . — doi : . — .
- Lanyi J. K. Bacteriorhodopsin (англ.) // Annu. Rev. Physiol. . — 2004. — Vol. 66 . — P. 665—688 . — doi : . — .
- Galagan J. E., Nusbaum C., Roy A., ; Allen, N; Naylor, J; Stange-Thomann, N; Dearellano, K; Johnson, R; Linton, L; Mcewan, P; Mckernan, K; Talamas, J; Tirrell, A; Ye, W; Zimmer, A; Barber, RD; Cann, I; Graham, DE; Grahame, DA; Guss, AM; Hedderich, R; Ingram-Smith, C; Kuettner, HC; Krzycki, JA; Leigh, JA; Li, W; Liu, J; Mukhopadhyay, B; Reeve, JN; Smith, K; Springer, TA; Umayam, LA; White, O; White, RH; Conway De Macario, E; Ferry, JG; Jarrell, KF; Jing, H; Macario, AJ; Paulsen, I; Pritchett, M; Sowers, KR; Swanson, RV; Zinder, SH; Lander, E; Metcalf, WW; Birren, B. et al. The genome of M. acetivorans reveals extensive metabolic and physiological diversity (англ.) // : journal. — 2002. — April ( vol. 12 , no. 4 ). — P. 532—542 . — doi : . — . — PMC .
- Waters E., ; Lin, X; Mathur, E; Ni, J; Podar, M; Richardson, T; Sutton, GG; Simon, M; Soll, D; Stetter, KO; Short, JM; Noordewier, M. et al. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2003. — Vol. 100 , no. 22 . — P. 12984—12988 . — doi : . — . — . — PMC . 16 октября 2019 года.
- Schleper C., Holz I., Janekovic D., Murphy J., Zillig W. (англ.) // : journal. — 1995. — 1 August ( vol. 177 , no. 15 ). — P. 4417—4426 . — . — PMC . 29 мая 2012 года.
- Sota M; Top E. M. // Plasmids: Current Research and Future Trends (англ.) . — , 2008.
- Xiang X., Chen L., Huang X., Luo Y., She Q., Huang L. (англ.) // : journal. — 2005. — Vol. 79 , no. 14 . — P. 8677—8686 . — doi : . — . — PMC . 16 октября 2019 года.
- Prangishvili D., Forterre P., Garrett R. A. Viruses of the Archaea: a unifying view (англ.) // Nat. Rev. Microbiol. : journal. — 2006. — Vol. 4 , no. 11 . — P. 837—848 . — doi : . — .
- Prangishvili D., Garrett R. A. (англ.) // : journal. — 2004. — Vol. 32 , no. Pt 2 . — P. 204—208 . — doi : . — . 18 апреля 2006 года.
- Pietilä M. K., Roine E., Paulin L., Kalkkinen N., Bamford D. H. An ssDNA virus infecting archaea; A new lineage of viruses with a membrane envelope (англ.) // : journal. — , 2009. — March ( vol. 72 , no. 2 ). — P. 307—319 . — doi : . — .
- Mojica F. J., Díez-Villaseñor C., García-Martínez J., Soria E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements (англ.) // : journal. — 2005. — Vol. 60 , no. 2 . — P. 174—182 . — doi : . — .
- Makarova K. S., Grishin N. V., Shabalina S. A., Wolf Y. I., Koonin E. V. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action (англ.) // : journal. — 2006. — Vol. 1 . — P. 7 . — doi : . — . — PMC .
- Graham D. E., Overbeek R., Olsen G. J., Woese C. R. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2000. — Vol. 97 , no. 7 . — P. 3304—3308 . — doi : . — . — . — PMC .
- ↑ Gaasterland T. Archaeal genomics (англ.) // Curr. Opin. Microbiol.. — 1999. — Vol. 2 , no. 5 . — P. 542—547 . — doi : . — .
- Allers T., Mevarech M. Archaeal genetics — the third way (англ.) // Nat. Rev. Genet. : journal. — 2005. — Vol. 6 , no. 1 . — P. 58—73 . — doi : . — .
- Werner F. Structure and function of archaeal RNA polymerases (англ.) // : journal. — , 2007. — September ( vol. 65 , no. 6 ). — P. 1395—1404 . — doi : . — .
- Aravind L., Koonin E. V. (англ.) // : journal. — 1999. — Vol. 27 , no. 23 . — P. 4658—4670 . — doi : . — . — PMC . 16 октября 2019 года.
- Lykke-Andersen J., Aagaard C., Semionenkov M., Garrett R. A. Archaeal introns: splicing, intercellular mobility and evolution (англ.) // : journal. — 1997. — September ( vol. 22 , no. 9 ). — P. 326—331 . — doi : . — .
- Watanabe Y., Yokobori S., Inaba T., et al. Introns in protein-coding genes in Archaea (англ.) // : journal. — 2002. — January ( vol. 510 , no. 1—2 ). — P. 27—30 . — doi : . — .
- Yoshinari S., Itoh T., Hallam S. J., et al. Archaeal pre-mRNA splicing: a connection to hetero-oligomeric splicing endonuclease (англ.) // : journal. — 2006. — August ( vol. 346 , no. 3 ). — P. 1024—1032 . — doi : . — .
- ↑ Bernander R. Archaea and the cell cycle (англ.) // : journal. — , 1998. — Vol. 29 , no. 4 . — P. 955—961 . — doi : . — .
- Kelman L. M., Kelman Z. Multiple origins of replication in archaea (англ.) // : journal. — 2004. — Vol. 12 , no. 9 . — P. 399—401 . — doi : . — .
- Onyenwoke R. U., Brill J. A., Farahi K., Wiegel J. Sporulation genes in members of the low G+C Gram-type-positive phylogenetic branch ( Firmicutes) (англ.) // : journal. — 2004. — Vol. 182 , no. 2—3 . — P. 182—192 . — doi : . — .
- Kostrikina N. A., Zvyagintseva I. S., Duda V. I. Cytological peculiarities of some extremely halophilic soil archaeobacteria (англ.) // : journal. — 1991. — Vol. 156 , no. 5 . — P. 344—349 . — doi : .
- DeLong E. F., Pace N. R. (англ.) // Syst. Biol. : journal. — 2001. — Vol. 50 , no. 4 . — P. 470—478 . — doi : . — .
- ↑ Pikuta E. V., Hoover R. B., Tang J. Microbial extremophiles at the limits of life (англ.) // : journal. — 2007. — Vol. 33 , no. 3 . — P. 183—209 . — doi : . — .
- Madigan M. T., Martino J. M. Brock Biology of Microorganisms (англ.) . — 11th. — Pearson, 2006. — P. 136. — ISBN 0-13-196893-9 .
- Takai K., Nakamura K., Toki T., Tsunogai U., Miyazaki M., Miyazaki J., Hirayama H., Nakagawa S., Nunoura T., Horikoshi K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — Vol. 105 , no. 31 . — P. 10949—10954 . — doi : . — . — . — PMC .
- Ciaramella M., Napoli A., Rossi M. Another extreme genome: how to live at pH 0 (англ.) // : journal. — 2005. — February ( vol. 13 , no. 2 ). — P. 49—51 . — doi : . — .
- Javaux E. J. Extreme life on Earth—past, present and possibly beyond (англ.) // Res. Microbiol. : journal. — 2006. — Vol. 157 , no. 1 . — P. 37—48 . — doi : . — .
- Nealson K. H. (англ.) // Orig Life Evol Biosph : journal. — 1999. — January ( vol. 29 , no. 1 ). — P. 73—93 . — doi : . — . 16 октября 2019 года.
- Davies P. C. The transfer of viable microorganisms between planets (англ.) // Ciba Found. Symp. : journal. — 1996. — Vol. 202 . — P. 304—314 . — .
- López-García P., López-López A., Moreira D., Rodríguez-Valera F. Diversity of free-living prokaryotes from a deep-sea site at the Antarctic Polar Front (англ.) // : journal. — 2001. — July ( vol. 36 , no. 2—3 ). — P. 193—202 . — .
- Karner M. B., DeLong E. F., Karl D. M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean (англ.) // Nature : journal. — 2001. — Vol. 409 , no. 6819 . — P. 507—510 . — doi : . — .
- Giovannoni S. J., Stingl U. Molecular diversity and ecology of microbial plankton (англ.) // Nature : journal. — 2005. — Vol. 427 , no. 7057 . — P. 343—348 . — doi : . — . — .
- DeLong E. F., Karl D. M. Genomic perspectives in microbial oceanography (англ.) // Nature. — 2005. — September ( vol. 437 , no. 7057 ). — P. 336—342 . — doi : . — . — .
- Konneke M., Bernhard A. E., de la Torre J. R., Walker C. B., Waterbury J. B., Stahl DA. Isolation of an autotrophic ammonia-oxidizing marine archaeon (англ.) // Nature : journal. — 2005. — Vol. 437 , no. 7057 . — P. 543—546 . — doi : . — . — .
- Agogué H., Maaike B., Dinasquet J., Herndl GJ.; Agogué, H; Brink, M; Dinasquet, J; Herndl, G.J. Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic (англ.) // Nature : journal. — 2008. — Vol. 456 , no. 7223 . — P. 788—791 . — doi : . — . — .
- Teske A., Sørensen K. B. Uncultured archaea in deep marine subsurface sediments: have we caught them all? (англ.) // : journal. — 2008. — January ( vol. 2 , no. 1 ). — P. 3—18 . — doi : . — .
- Lipp J. S., Morono Y., Inagaki F., Hinrichs K. U. Significant contribution of Archaea to extant biomass in marine subsurface sediments (англ.) // Nature : journal. — 2008. — July ( vol. 454 , no. 7207 ). — P. 991—994 . — doi : . — . — .
- Cabello P., Roldán M. D., Moreno-Vivián C. (англ.) // : journal. — , 2004. — November ( vol. 150 , no. Pt 11 ). — P. 3527—3546 . — doi : . — . 7 июня 2011 года.
- Mehta M. P., Baross J. A. Nitrogen fixation at 92 degrees C by a hydrothermal vent archaeon (англ.) // Science : journal. — 2006. — December ( vol. 314 , no. 5806 ). — P. 1783—1786 . — doi : . — . — .
- Coolen M. J., Abbas B., van Bleijswijk J., et al. Putative ammonia-oxidizing Crenarchaeota in suboxic waters of the Black Sea: a basin-wide ecological study using 16S ribosomal and functional genes and membrane lipids (англ.) // Environ. Microbiol. : journal. — 2007. — April ( vol. 9 , no. 4 ). — P. 1001—1016 . — doi : . — .
- Leininger S., Urich T., Schloter M., et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils (англ.) // Nature : journal. — 2006. — August ( vol. 442 , no. 7104 ). — P. 806—809 . — doi : . — . — .
- Baker, B. J.; Banfield, J. F. Microbial communities in acid mine drainage (англ.) // : journal. — 2003. — Vol. 44 , no. 2 . — P. 139—152 . — doi : . — . (недоступная ссылка)
- Schimel J. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2004. — August ( vol. 101 , no. 34 ). — P. 12400—12401 . — doi : . — . — . — PMC . 16 октября 2019 года.
- . Дата обращения: 26 июня 2008. 21 мая 2008 года.
- (23 апреля 2008). Дата обращения: 26 июня 2008. 14 мая 2008 года.
- . Climate Change 2001 . United Nations Environment Programme. 5 августа 2012 года.
- Eckburg P., Lepp P., Relman D. Archaea and their potential role in human disease (англ.) // : journal. — 2003. — Vol. 71 , no. 2 . — P. 591—596 . — doi : . — . — PMC .
- Cavicchioli R., Curmi P., Saunders N., Thomas T. Pathogenic archaea: do they exist? (англ.) // : journal. — 2003. — Vol. 25 , no. 11 . — P. 1119—1128 . — doi : . — .
- Lepp P., Brinig M., Ouverney C., Palm K., Armitage G., Relman D. Methanogenic Archaea and human periodontal disease (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2004. — Vol. 101 , no. 16 . — P. 6176—6181 . — doi : . — . — . — PMC .
- Vianna M. E., Conrads G., Gomes B. P., Horz H. P. (англ.) // J. Clin. Microbiol. : journal. — 2006. — April ( vol. 44 , no. 4 ). — P. 1274—1282 . — doi : . — . — PMC . 16 октября 2019 года.
- Waters E., Hohn M. J., Ahel I., ; Lin, X; Mathur, E; Ni, J; Podar, M; Richardson, T; Sutton, GG; Simon, M; Soll, D; Stetter, KO; Short, JM; Noordewier, M. et al. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2003. — October ( vol. 100 , no. 22 ). — P. 12984—12988 . — doi : . — . — . — PMC . 16 октября 2019 года.
- Jahn U., Gallenberger M., Paper W., et al. (англ.) // : journal. — 2008. — March ( vol. 190 , no. 5 ). — P. 1743—1750 . — doi : . — . — PMC . 27 мая 2020 года.
- ↑ рус. Архееподобные ацидофильные наноорганизмы Ричмондских рудников
- Baker B. J., Comolli L. R., Dick G. J., Hauser L. J., Hyatt D., Dill B. D., Land M. L., VerBerkmoes N. C., Hettich R. L., Banfield J. F. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2010. — May ( vol. 107 , no. 19 ). — P. 8806—8811 . — doi : . — . — PMC . 10 февраля 2021 года.
- Chaban B., Ng S. Y., Jarrell K. F. Archaeal habitats—from the extreme to the ordinary (англ.) // : journal. — , 2006. — February ( vol. 52 , no. 2 ). — P. 73—116 . — doi : . — .
- Schink B. Energetics of syntrophic cooperation in methanogenic degradation (англ.) // : journal. — , 1997. — June ( vol. 61 , no. 2 ). — P. 262—280 . — . — PMC .
- Lange, M; Lange, M.; Westermann, P; Westermann, P.; Ahring, BK; Ahring, B. K. Archaea in protozoa and metazoa (англ.) // : journal. — Springer , 2005. — Vol. 66 , no. 5 . — P. 465—474 . — doi : . — .
- van Hoek A. H., van Alen T. A., Sprakel V. S., et al. (англ.) // : journal. — Oxford University Press , 2000. — 1 February ( vol. 17 , no. 2 ). — P. 251—258 . — . 16 октября 2019 года.
- Preston, C. M.; Wu, K. Y.; Molinski, T. F.; Delong, E. F. A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1996. — Vol. 93 , no. 13 . — P. 6241—6246 . — doi : . — . — . — PMC .
- Eckburg P. B., Bik E. M., Bernstein C. N., et al. Diversity of the human intestinal microbial flora (англ.) // Science. — 2005. — June ( vol. 308 , no. 5728 ). — P. 1635—1638 . — doi : . — . — . — PMC .
- Samuel B. S., Gordon J. I. A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2006. — June ( vol. 103 , no. 26 ). — P. 10011—10016 . — doi : . — . — . — PMC .
- Wegley, L; Wegley, L.; Yu; Yu, Y.; Breitbart; Breitbart, M.; Casas; Casas, V.; Kline; Kline, D. I.; Rohwer; Rohwer, F. (англ.) // : journal. — 2004. — Vol. 273 . — P. 89—96 . — doi : . 11 сентября 2008 года. . Дата обращения: 21 июля 2012. Архивировано из 11 сентября 2008 года.
- Chelius M. K., Triplett E. W. (англ.) // Microb. Ecol. : journal. — 2001. — April ( vol. 41 , no. 3 ). — P. 252—263 . — doi : . — .
- Simon H. M., Dodsworth J. A., Goodman R. M. Crenarchaeota colonize terrestrial plant roots (англ.) // Environ. Microbiol.. — 2000. — October ( vol. 2 , no. 5 ). — P. 495—505 . — doi : . — .
- Gevers D., Dawyndt P., Vandamme P., et al. (англ.) // Philos. Trans. R. Soc. Lond., B, Biol. Sci. : journal. — 2006. — Vol. 361 , no. 1475 . — P. 1911—1916 . — doi : . — . — PMC . 20 декабря 2012 года.
- ↑ Robertson C. E., Harris J. K., Spear J. R., Pace N. R. Phylogenetic diversity and ecology of environmental Archaea (англ.) // Curr. Opin. Microbiol. : journal. — 2005. — Vol. 8 , no. 6 . — P. 638—642 . — doi : . — .
- Huber H., Hohn M. J., Rachel R., Fuchs T., Wimmer V. C., Stetter K. O. A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont (англ.) // Nature : journal. — 2002. — Vol. 417 , no. 6884 . — P. 27—28 . — doi : . — .
- Barns S. M., Delwiche C. F., Palmer J. D., Pace N. R. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1996. — Vol. 93 , no. 17 . — P. 9188—9193 . — doi : . — . — . — PMC . 8 мая 2020 года.
- Elkins J. G., Podar M., Graham D. E., ; Goltsman, E; Barry, K; Koonin, EV; Hugenholtz, P; Kyrpides, N; Wanner, G; Richardson, P; Keller, M; Stetter, K.O. et al. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2008. — June ( vol. 105 , no. 23 ). — P. 8102—8107 . — doi : . — . — . — PMC . 27 мая 2020 года.
- Baker B. J., Tyson G. W., Webb R. I., Flanagan J., Hugenholtz P. and Banfield J. F. Lineages of acidophilic Archaea revealed by community genomic analysis. Science (англ.) // Science : journal. — 2006. — Vol. 314 , no. 6884 . — P. 1933—1935 . — doi : . — . — .
- Baker B. J., Comolli L. R., Dick G. J., et al. Enigmatic, ultrasmall, uncultivated Archaea (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2010. — May ( vol. 107 , no. 19 ). — P. 8806—8811 . — doi : . — . — . — PMC .
- de Queiroz K. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2005. — Vol. 102 , no. Suppl 1 . — P. 6600—6607 . — doi : . — . — . — PMC . 16 октября 2019 года.
- Eppley J. M., Tyson G. W., Getz W. M., Banfield J. F. (англ.) // Genetics : journal. — 2007. — Vol. 177 , no. 1 . — P. 407—416 . — doi : . — . — PMC . 16 октября 2019 года.
- Papke R. T., Zhaxybayeva O., Feil E. J., Sommerfeld K., Muise D., Doolittle W. F. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2007. — Vol. 104 , no. 35 . — P. 14092—14097 . — doi : . — . — . — PMC . 16 октября 2019 года.
- Kunin V., Goldovsky L., Darzentas N., Ouzounis C. A. (англ.) // : journal. — 2005. — Vol. 15 , no. 7 . — P. 954—959 . — doi : . — . — PMC . 16 октября 2019 года.
- Hugenholtz P. (англ.) // . — 2002. — Vol. 3 , no. 2 . — P. REVIEWS0003 . — doi : . — . — PMC . 15 марта 2020 года.
- Rappé M. S., Giovannoni S. J. The uncultured microbial majority (англ.) // Annu. Rev. Microbiol. . — 2003. — Vol. 57 . — P. 369—394 . — doi : . — .
- Морозова О. В. Загадки архей и их фагов // Вестник ВОГиС. — 2005. — Т. 9 , № 1 . — С. 55—66 .
- Spang A., Hatzenpichler R., Brochier-Armanet C., Rattei T., Tischler P., Spieck E., Streit W., Stahl D. A., Wagner M., Schleper C. (англ.) // : journal. — 2010. — Vol. 18 , no. 8 . — P. 331—340 . — . 2 июня 2013 года.
- ↑ Spang A. , Saw J. H. , Jørgensen S. L. , Zaremba-Niedzwiedzka K. , Martijn J. , Lind A. E. , van Eijk R. , Schleper C. , Guy L. , Ettema T. J. (англ.) // Nature. — 2015. — doi : . — .
- Seitz K. W. , Lazar C. S. , Hinrichs K. U. , Teske A. P. , Baker B. J. (англ.) // The ISME journal. — 2016. — Vol. 10, no. 7 . — P. 1696—1705. — doi : . — .
- Breithaupt H. The hunt for living gold. The search for organisms in extreme environments yields useful enzymes for industry (англ.) // : journal. — 2001. — Vol. 2 , no. 11 . — P. 968—971 . — doi : . — . — PMC .
- ↑ Egorova K., Antranikian G. Industrial relevance of thermophilic Archaea (англ.) // Curr. Opin. Microbiol.. — 2005. — Vol. 8 , no. 6 . — P. 649—655 . — doi : . — .
- Synowiecki J., Grzybowska B., Zdziebło A. Sources, properties and suitability of new thermostable enzymes in food processing (англ.) // : journal. — 2006. — Vol. 46 , no. 3 . — P. 197—205 . — doi : . — .
- Jenney F. E., Adams M. W. The impact of extremophiles on structural genomics (and vice versa) (англ.) // Extremophiles : journal. — 2008. — January ( vol. 12 , no. 1 ). — P. 39—50 . — doi : . — .
- Schiraldi C., Giuliano M., De Rosa M. (англ.) // Archaea : journal. — 2002. — Vol. 1 , no. 2 . — P. 75—86 . — doi : . — . — PMC . 26 августа 2013 года. . Дата обращения: 21 июля 2012. Архивировано из 26 августа 2013 года.
- Norris P. R., Burton N. P., Foulis N. A. Acidophiles in bioreactor mineral processing (англ.) // Extremophiles. — 2000. — Vol. 4 , no. 2 . — P. 71—6 . — doi : . — .
- O'Connor E. M., Shand R. F. Halocins and sulfolobicins: the emerging story of archaeal protein and peptide antibiotics (англ.) // J. Ind. Microbiol. Biotechnol. : journal. — 2002. — January ( vol. 28 , no. 1 ). — P. 23—31 . — doi : . — .
- Shand R. F.; Leyva K. J. Archaeal Antimicrobials: An Undiscovered Country // Archaea: New Models for Prokaryotic Biology (англ.) / Blum P (ed.). — , 2008. — ISBN 978-1-904455-27-1 .
Литература
- Воробьева Л. В. Археи: Учебное пособие для вузов. — М.: Академкнига, 2007. — 447 с.
- Громов Б. В. // СОЖ. — 1997. — № 4 . — С. 23—26 .
- Морозова О. В. Загадки архей и их фагов // Вестник ВОГиС. — 2005. — Том 9. — № 1. — С. 55—66
- Thomas Cavalier-Smith. . — 2006.
Ссылки
- Волкова, Ольга. // Сайт Biomolecula.ru (22 января 2015). Дата обращения: 5 апреля 2018.
- Старокадомский, Пётр. . // Сайт Biomolecula.ru (7 февраля 2013). Дата обращения: 5 апреля 2018.
- Панов, Андрей. . // Сайт Biomolecula.ru (22 сентября 2015). Дата обращения: 5 апреля 2018.
- Конышев, Илья. . // Сайт Biomolecula.ru (3 сентября 2015). Дата обращения: 5 апреля 2018.
- Кондратенко, Юлия. . // Сайт Biomolecula.ru (22 декабря 2015). Дата обращения: 5 апреля 2018.
|
Классификация
|
|
|---|---|
| Euryarchaeota | |
|
|
|
|
Эта статья входит в число
избранных статей
русскоязычного раздела Википедии.
|
- 2020-02-08
- 1