Interested Article - Иммунная система
- 2020-10-30
- 1

Имму́нная систе́ма — система биологических структур и процессов организма, обеспечивающая его защиту от инфекций , токсинов и злокачественных клеток . Для правильной работы иммунной системы необходимо, чтобы она умела распознавать широкий спектр патогенов — от вирусов до многоклеточных червей — и отличать их от собственных здоровых тканей организма . У многих видов имеются две подсистемы: врождённая иммунная система и приобретённая (адаптивная) иммунная система. Обе подсистемы используют как гуморальные механизмы, так и клеточные механизмы. К числу древнейших механизмов иммунной системы животных относят фагоцитоз , систему комплемента и антимикробные пептиды . Адаптивная иммунная система в ходе эволюции появилась у челюстноротых позвоночных животных . Один из важнейших механизмов адаптивной иммунной системы — иммунологическая память , благодаря которой организм развивает более сильный иммунный ответ на патоген после первой встречи с ним. Основу вакцинации составляет именно иммунологическая память.
Нарушения в работе иммунной системы приводят к возникновению аутоиммунных заболеваний , воспалительных заболеваний и рака . Когда иммунная система функционирует слабо, наблюдаются иммунодефицитные состояния, из-за которых организм становится более уязвим для инфекций. Иммунодефицит может быть как врождённым, обусловленным генетическими отклонениями, так и приобретённым, например, в результате ВИЧ-инфекции или приёма препаратов-иммуносупрессоров .
Функционирование иммунной системы изучает наука иммунология .
Органы иммунной системы
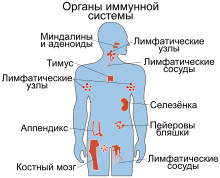
Иммунную систему образуют органы и анатомические структуры, содержащие лимфоидную ткань и участвующие в образовании клеток, которые осуществляют защитную реакцию организма, создавая иммунитет . У человека с 13-й недели после зачатия некоторые стволовые клетки перемещаются в тимус и костный мозг , которые называют первичными (центральными) лимфоидными органами . Все клетки крови , включая лейкоциты , происходят из общего предшественника — плюрипотентной стволовой клетки костного мозга . Лимфоциты — T-лимфоциты (T-клетки), B-лимфоциты (B-клетки), NK-клетки — обособляются в отдельные клеточные линии на ранних этапах, а гранулоциты некоторое время развиваются вместе с эритроцитами и тромбоцитами . К числу гранулоцитов, выполняющих защитные функции, относят базофилы , эозинофилы , нейтрофилы , циркулирующие в кровотоке, и макрофаги , локализованные в тканях. Особую группу макрофагов составляют микроглиальные клетки, на которые приходится от 10 % до 15 % клеток головного мозга . Клетки микроглии поглощают инфекционные агенты и в нервной ткани головного и спинного мозга , уничтожают повреждённые нейроны и ненужные синапсы . T-лимфоциты обязательно проходят стадию созревания в тимусе, а B-лимфоциты полностью созревают в костном мозге. Созревшие T- и B-клетки покидают места дифференцировки и перемещаются во вторичные (периферические) лимфоидные органы — селезёнку , лимфатические узлы , ( ) и другие органы .
Уровни защиты
Иммунная система обеспечивает защиту организма от инфекций на нескольких уровнях с повышающейся специфичностью. Организм имеет физические барьеры, мешающие проникновению в него вирусов и бактерий . Если патогену удаётся их преодолеть, то он сталкивается со врождённой иммунной системой, которая обеспечивает быстрый, но неспецифический ответ. Врождённая иммунная система имеется у растений и животных . У позвоночных, если патоген преодолевает врождённый иммунный ответ, он сталкивается со следующим рубежом защиты — адаптивной иммунной системой. Адаптивная иммунная система обеспечивает специфический иммунный ответ, направленный против конкретного патогена. После того, как патоген был уничтожен, адаптивная иммунная система «запоминает» его с помощью иммунологической памяти, благодаря которой при повторной встрече с патогеном организм сможет быстро развить специфический иммунный ответ против него . В таблице ниже перечислены основные компоненты врождённой и адаптивной иммунной систем.
| Врождённая иммунная система | Адаптивная иммунная система |
|---|---|
| Ответ неспецифичен | Специфический ответ |
| Обеспечивает немедленный иммунный ответ умеренной силы | Обеспечивает отложенный сильный иммунный ответ |
| Имеет клеточную и гуморальную составляющую | Имеет клеточную и гуморальную составляющую |
| Нет иммунологической памяти | После первого столкновения с патогеном появляется иммунологическая память |
| Есть практически у всех форм жизни | Есть только у челюстноротых позвоночных животных |
И врождённая, и адаптивная иммунная системы полагаются на способность организма отличать своё от не-своего. В иммунологии под «своим» понимают молекулы , которые являются компонентами нормального здорового организма. «Не-свои» молекулы — это молекулы чужеродного происхождения, которые отсутствуют у здорового организма. Часть чужеродных молекул может связываться с особыми рецепторами на иммунных клетках и запускать иммунный ответ; такие молекулы называют антигенами .
Новорождённые особенно уязвимы для инфекций, поскольку до рождения они не сталкивались с патогенами. Несколько уровней защиты новорождённый получает от своей матери. В ходе беременности антитела класса G ( иммуноглобулины G , IgG) передаются от матери к плоду непосредственно через плаценту , поэтому у людей новорождённый имеет высокий уровень антител с рождения, которые соответствуют тем антигенам, с которыми сталкивалась его мать . Грудное молоко или молозиво также содержат антитела, которые также обеспечивают защиту новорождённого, пока его собственная иммунная система не начнёт вырабатывать свои антитела . Поскольку организм новорождённого сам не вырабатывает антител и не формирует клетки памяти, его иммунитет является пассивным. Пассивная защита новорождённого, как правило, недолговечна и функционирует от нескольких дней до нескольких недель. Пассивный иммунитет можно создать и искусственно путём введения в организм сыворотки , богатой антителами .
Поверхностные барьеры
Организм защищён от проникновения патогенов несколькими поверхностными барьерами: механическими, химическими и биологическими. Восковая кутикула , покрывающая листья растений, экзоскелет насекомых , раковины, скорлупа яиц и кожа могут служить примерами механических барьеров, которые являются первым препятствием на пути проникновения инфекции в организм . Однако, так как организм не может быть полностью изолированным от окружающей среды, некоторые органы, непосредственно контактирующие со внешней средой и её компонентами — лёгкие , желудочно-кишечный тракт и мочеполовая система — нуждаются в дополнительном уровне защиты. От внешней среды организм животного защищён эпителием , который выстилает пищеварительную систему, дыхательные пути и мочеполовой тракт. Эпителий, выстилающий сообщающиеся со внешней средой органы, называют слизистой оболочкой. Клетки эпителия выделяют разнообразные соединения, убивающие бактерий или подавляющие их рост (подробнее о защитных веществах эпителия см. следующий абзац). Помимо локальных химических факторов, в слизистых присутствуют фагоцитирующие клетки (они особенно многочисленны в слизистых лёгких) . Благодаря кашлю и чиханию патогены механически удаляются из дыхательных путей . Слёзы и моча обеспечивают вымывание патогенов, а слизь , выделяемая клетками дыхательных путей и желудочно-кишечного тракта, склеивает и обездвиживает клетки патогенов .
К числу химических барьеров относят антимикробные пептиды , такие как β- дефензины , которые секретируются клетками дыхательных путей и кожи . Антимикробные белки , такие как лизоцим и фосфолипаза A2 , содержатся в слюне, слезах и грудном молоке . После менархе вагина начинает выделять вещества, обеспечивающие химический барьер за счёт закисления среды. В сперме содержатся дефензины и ионы цинка , обладающие антимикробным действием . В желудке мощную защиту от поглощённых с пищей патогенов обеспечивает соляная кислота , секретируемая некоторыми клетками стенки желудка .
В пищеварительной и мочеполовой системах имеется своя комменсальная микрофлора , которая служит биологическим барьером, поскольку за счёт конкурентных механизмов лишает патогенные микроорганизмы ресурсов для существования или делает окружающие условия неблагоприятными для них, закисляя среду . Таким образом, в этих органах имеет место симбиотическая связь микрофлоры и иммунной системы. Однако, так как большинство антибиотиков поражают бактерий неспецифически и наряду с патогенными бактериями убивают полезную микрофлору, после приёма антибиотиков освободившееся пространство могут колонизировать грибы , например, дрожжи Candida , вызывающие кандидоз . Поэтому после приёма антибиотиков с целью восстановления нормальной микрофлоры назначают препараты групп пребиотиков и пробиотиков . Здоровая микрофлора обеспечивает мощную защиту от бактериального гастроэнтерита , , инфекций мочеполовой системы и бактериальных осложнений после хирургических вмешательств .
Врождённая иммунная система
Микроорганизмы и токсины, успешно преодолевшие физические защитные барьеры организма, сталкиваются с противодействием врождённой иммунной системы. Врождённый иммунный ответ, как правило, запускается после распознавания патогенов рецепторами опознавания патогенов , узнающими молекулы, имеющиеся у большинства представителей больших групп патогенов (как липополисахарид у грамотрицательных бактерий ) . Врождённый иммунный ответ также запускается при распознавании рецепторами сигналов, исходящих от повреждённых клеток организма или клеток, находящихся в состоянии . Врождённая иммунная система неспецифична и обеспечивает ответ на широкий спектр патогенов независимо от их специфических свойств . Врождённый иммунитет не обеспечивает долговременную защиту от патогена, тем не менее, эта форма иммунитета доминирует у большинства организмов .
Распознавание паттернов

Клетки врождённой иммунной системы распознают молекулы и молекулярные комплексы, продуцируемые микробными клетками, с помощью рецепторов опознавания паттернов ( англ. pattern recognition receptors, PRRs ) . Гены , кодирующие PRRs, сохраняются в неизменном виде и не претерпевают перестроек в течение жизни организма. PRRs экспрессируются преимущественно клетками врождённого иммунитета, в частности, дендритными клетками , макрофагами , нейтрофилами , а также эпителиальными клетками. PRPs распознают молекулярные паттерны двух классов: ( англ. pathogen-associated molecular patterns, PAMPs ), и молекулярные паттерны, ассоциированные с повреждениями ( англ. damage-associated molecular patterns, DAMPs ). PAMPs узнают клетки патогенов, а DAMPs узнают клетки самого организма, подвергшиеся стрессу или повреждениям .
Распознавание внеклеточных или заключённых в эндосомы PAMPs осуществляют трансмембранные белки , известные как Toll-подобные рецепторы ( англ. Toll-like receptors, TLRs ) . TLRs имеют характерный , известный как ( англ. Leucine rich repeats (LRR) ), которые играют важную роль как в поддержании структуры TLRs, так и их функционировании . Первые TLRs были описаны у плодовой мушки Drosophila . TLRs запускают синтез и секрецию цитокинов , которые активируют защитные программы врождённого или адаптивного иммунитета. У человека описано 10 функциональных TLR .
Клетки врождённой иммунной системы имеют рецепторы, которые распознают опасные молекулярные паттерны, свидетельствующие об инфекции или повреждении клетки, в цитозоле . Среди этих рецепторов выделяют три ключевых класса: NOD-подобные рецепторы , RIG-подобные рецепторы и цитозольные сенсоры ДНК . В ответ на цитозольные PAMPs и DAMPs собираются инфламмасомы — мультибелковые комплексы , эффекторным компонентом которых является каспаза 1 . Инфламмасомы обеспечивают продукцию активных форм воспалительных цитокинов IL-1β и IL-18 .
Клеточные компоненты

Важнейшую роль в функционировании врождённого иммунитета играют лейкоциты . Среди лейкоцитов врождённого иммунитета выделяют фагоциты (макрофаги, нейтрофилы, дендритные клетки), , тучные клетки , базофилы , эозинофилы и натуральные киллеры . Эти клетки распознают клетки патогенов и убивают их . Клетки врождённого иммунитета играют важную роль в развитии лимфоидных органов и активации адаптивного иммунитета .
Многие клетки врождённого иммунитета обладают способностью к фагоцитозу, то есть поглощению, патогенных агентов. Фагоциты «патрулируют» организм в поисках патогенных клеток или же направленно мигрируют к очагу инфекции по направлению, указываемому градиентом концентрации цитокинов . Когда фагоцит поглощает патогенную клетку, она помещается внутрь особой везикулы — фагосомы , которая далее сливается с лизосомой с образованием . Внутри фаголизосомы патогенная клетка погибает под действием расщепляющих ферментов или свободных радикалов , образующихся в результате . Фагоцитоз, возможно, является древнейшим защитным механизмом, поскольку фагоциты имеются и у хордовых , и у беспозвоночных животных .
Крупнейшие группы фагоцитов — нейтрофилы и макрофаги . Нейтрофилы в норме циркулируют по кровотоку и являются самой многочисленной группой фагоцитов, составляя от 50 % до 60 % всех лейкоцитов в крови . В ходе острой фазы воспаления нейтрофилы мигрируют к очагу воспаления посредством хемотаксиса и, как правило, первыми из иммунных клеток прибывают в очаг инфекции. Макрофаги, в отличие от нейтрофилов, находятся в тканях и не циркулируют по кровеносным сосудам . Макрофаги секретируют разнообразные вещества, такие как ферменты, белки системы комплемента и цитокины, поглощают остатки погибших клеток организма, а также выступают в роли антигенпрезентирующих клеток , активирующих адаптивный иммунный ответ .
В тканях, контактирующих со внешней средой, находятся фагоциты, относящиеся к числу дендритных клеток. Дендритные клетки обнаруживаются в коже, эпителии ноздрей , лёгких, желудка и кишечника. Своё название дендритные клетки получили из-за характерных отростков, напоминающих дендриты нейронов , однако никакого отношения к нервной системе они не имеют. Дендритные клетки служат связующим звеном между тканями тела и иммунной системы благодаря способности к презентации антигенов T-лимфоцитам (T-клеткам) .
Лейкоциты, в цитоплазме которых содержатся гранулы, получили общее название гранулоциты. К гранулоцитам относятся нейтрофилы, базофилы и эозинофилы. В соединительных тканях и слизистых оболочках находятся тучные клетки, содержащие гранулы в цитоплазме и регулирующие воспалительный ответ . Тучные клетки играют важную роль в развитии аллергических реакций и анафилаксии . Эозинофилы и базофилы секретируют вещества, направленные на борьбу с многоклеточными паразитами , а также задействованы в аллергических реакциях .
Врождённые лимфоидные клетки являются производными общей клетки-предшественницы иммунных клеток. Они не имеют миелоидных или дендритных клеток, а также не несут T - и B-клеточных рецепторов . К числу врождённых лимфоидных клеток относятся натуральные киллеры (NK-клетки), которые не уничтожают инфекционные агенты непосредственно, а убивают клетки организма, инфицированные вирусом или внутриклеточной бактерией, и злокачественные клетки . Как правило, инфицированные клетки не имеют на поверхности молекул главного комплекса гистосовместимости (MHC) I класса ( ) , и NK-клетки уничтожают их без какой-либо предварительной активации. Нормальные клетки организма экспрессируют MHC-I и не становятся жертвами NK-клеток .
Воспаление
Воспаление является одной из первых реакций иммунной системы на развивающуюся инфекцию . К числу симптомов воспаления относят покраснение, отёк, повышение температуры и болезненность воспалённого участка, которые обусловлены повышенным кровотоком в очаге воспаления. Повреждённые и инфицированные клетки выделяют эйкозаноиды и цитокины, которые привлекают к очагу воспаления иммунные клетки. Одна из групп эйкозаноидов, простагландины , вызывают лихорадку и связанное с воспалением расширение кровеносных сосудов, а представители другой группы эйкозаноидов, лейкотриены , привлекают в очаг воспаления некоторые лейкоциты . К числу наиболее распространённых цитокинов относят интерлейкины , с помощью них лейкоциты «общаются» друг с другом, хемокины , стимулирующие хемотаксис, и интерфероны , обладающие противовирусными свойствами, так как подавляют синтез белков в инфицированной клетке . При воспалении также происходит высвобождение цитотоксических факторов и факторов роста . Все эти вещества привлекают иммунные клетки в очаг инфекции и способствуют заживлению повреждённой ткани после уничтожения патогенных клеток .
Система комплемента

Система комплемента представляет собой биохимический каскад , направленный на нарушение целостности чужеродных клеток. В состав системы комплемента входит более 20 белков, которые дополняют («комплементируют») действие антител по уничтожению патогенов . Система комплемента — важнейший гуморальный компонент врождённого иммунитета. Система комплемента есть не только у позвоночных, но и у беспозвоночных животных и даже растений .
У человека компоненты системы комплемента связываются либо с антителами, уже прикрепившимися к микробным клеткам, либо с углеводными фрагментами на поверхности микробных клеток. Связывание белков комплемента с мишенью запускает сигнальный каскад, на каждом этапе которого сигнал усиливается благодаря активации протеаз , входящих в состав системы комплемента, посредством протеолиза . Активированные протеазы далее за счёт протеолиза активируют новые протеазы, и так далее, то есть система комплемента активируется по механизму положительной обратной связи . В конечном счёте активация системы комплемента приводит к продукции пептидов, которые привлекают к патогену другие иммунные клетки, увеличивают проницаемость стенок кровеносных сосудов и опсонизируют (покрывают) патогенную клетку, помечая её для дальнейшего разрушения. Белки системы комплемента также могут непосредственно убивать микробные клетки, встраиваясь в их мембраны и нарушая их целостность .
Адаптивная иммунная система
В ходе эволюции адаптивная иммунная система появилась у челюстноротых животных. Адаптивный иммунитет обеспечивает сильный специфический ответ, а также формирование иммунологической памяти. Адаптивный иммунный ответ специфичен по отношению к определённому антигену, распознавание которого клетками адаптивного иммунитета происходит в ходе процесса презентации антигена. После уничтожения патогена сохраняются клетки адаптивного иммунитета, которые хранят сведения о его антигенах и обеспечивают иммунологическую память. Благодаря ей при вторичном проникновении патогена на него развивается быстрый специфический иммунный ответ .
Распознавание антигена
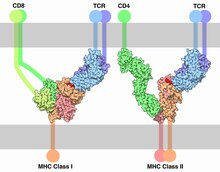
Клетки адаптивного иммунитета представлены специфической группой лейкоцитов — лимфоцитами, которые подразделяют на T-лимфоциты (T-клетки) и B-лимфоциты (B-клетки). Лимфоциты образуются от гемоцитобластов в костном мозге, и далее T-клетки созревают в тимусе, а B-клетки созревают в костном мозге. T-клетки обеспечивают клеточный адаптивный иммунный ответ, а B-клетки — гуморальный адаптивный иммунный ответ. T- и B-клетки несут на своей поверхности рецепторы (T- и B-клеточные рецепторы соответственно), распознающие антигены. Как правило, T-клетки не способны распознавать антигены в исходной форме; они распознают только фрагменты антигенов, связанные с молекулами MHC на поверхности антигенпрезентирующих клеток. T-киллеры распознают фрагменты антигенов, связанные с MHC-I, а T-хелперы и регуляторные T-клетки распознают фрагменты антигенов в комплексе с . Особая группа T-клеток, , могут распознавать интактные антигены, не связанные с MHC . B-клеточный рецептор представляет собой молекулу антитела, заякоренную на поверхности B-клетки, и распознаёт антиген без дополнительного процессинга. B-клеточные рецепторы разных линий B-клеток соответствуют разным антигенам и отображают весь репертуар антител, которые могут быть образованы .
Клеточный адаптивный иммунный ответ

Среди T-клеток выделяют три основные популяции: T-киллеры, T-хелперы и регуляторные T-клетки. T-киллеры уничтожают инфицированные и повреждённые клетки . Как в случае с B-клетками, T-клетки одной линии распознают один и тот же антиген. T-киллеры активируются, когда их T-клеточные рецепторы ( англ. T cell receptors, TCRs ) распознают фрагмент антигена в комплексе с MHC-I на поверхности антигенпрезентирующей клетки. Для распознавания такого комплекса T-киллерами, помимо TCRs, необходим корецептор CD8 . После активации T-киллер мигрирует по кровотоку в поисках такого же антигена, как тот, который был её представлен. Когда T-киллер встречает клетку с нужным антигеном, он высвобождает белки с цитотоксическими свойствами, такие как перфорины , которые встраиваются в мембрану клетки-мишени с образованием пор и нарушают её целостность. Проникновение в клетку-мишень другого белка, протеазы , запускает апоптоз клетки-мишени . T-киллеры играют особо важную роль в предотвращении репликации вирусов за счёт уничтожения инфицированных клеток. Активация T-киллеров жёстко регулируется и происходит только в случае почти идеального соответствия T-клеточного рецептора и антигена, кроме того, для активации T-киллеров необходимы дополнительные сигналы, которые посылаются T-клетками другого типа — T-хелперами .
T-хелперы регулируют адаптивный и приобретённый иммунные ответы. T-хелперы лишены цитотоксической активности, они не уничтожают ни клетки патогена, ни заражённые клетки . На поверхности T-хелперов имеются TCRs, которые распознают фрагменты антигенов в комплексе с MHC-II. Как и в случае T-киллеров, в дополнение к TCR для распознавания комплекса MHC:антиген T-хелперы используют корецептор, но не CD8, а CD4 , который запускает сигнальные каскады, активирующие T-хелпер. T-хелперы связываются с комплексом MHC:антиген слабее, чем T-киллеры, и для активации T-хелпера необходимо, чтобы 200—300 TCRs на его поверхности были связаны с комплексом MHC:антиген, в то время как активация T-киллера происходит уже тогда, когда с комплексом MHC:антиген связан всего один рецептор. Кроме того, для активации T-хелперов требуется более длительное взаимодействие с антигенпрезентирующей клеткой . Активированный T-хелпер высвобождает цитокины, воздействующие на клетки многих типов. Цитокиновые сигналы T-хелперов усиливают бактерицидные свойства макрофагов и активность T-киллеров .
Регуляторные T-клетки, ранее известные как супрессорные T-клетки, подавляют функционирование и пролиферацию эффекторных T-клеток, предотвращая развитие аутоиммунных заболеваний , и по происхождению родственны T-хелперам. Как и T-хелперы, регуляторные T-клетки экспрессируют корецептор CD4 . γδ-T клетки экспрессируют альтернативную форму TCR, отличающуюся от таковой у CD4+ и CD8+ T-клеток, и совмещают свойства T-хелперов, T-киллеров и NK-клеток .
Гуморальный адаптивный иммунный ответ
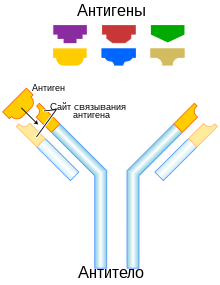
B-клетки распознают антигены посредством B-клеточных рецепторов, которые представляют собой антитела, заякоренные на поверхности B-клеток . Когда B-клеточный рецептор связывается с антигеном, он интернализуется, и внутри B-клетки антиген расщепляется на пептиды посредством протеолиза. Полученные фрагменты антигенов B-клетка экспонирует на своей поверхности в комплексе с MHC-II. Комплекс антиген: MHC-II активирует T-хелпер, который выделяет лимфокины , активирующие B-клетку . Активированная B-клетка начинает делиться, и её клетки-потомки, называемые плазматическими клетками , секретируют миллионы молекул антител, которые идентичны B-клеточному рецептору, первоначально связавшему антиген. Антитела циркулируют в крови и лимфе , связываются с клетками патогена, экспрессирующими соответствующий им антиген, и маркируют их для разрушения белками комплемента или фагоцитами. Антитела могут сами по себе обладать защитными свойствами, связываясь с бактериальными токсинами и нейтрализуя их или конкурируя с вирусами и бактериями за рецепторы, мешая им инфицировать клетку .
Физиологическая регуляция

Иммунная система тесно взаимодействует с другими системами органов, в частности, эндокринной и нервной . Иммунная система также играет важную роль в восстановлении целостности тканей и регенерации .
Гормоны
Гормоны действуют как иммуномодуляторы, регулирующие чувствительность иммунной системы. Например, женские половые гормоны являются иммуностимуляторами и для врождённого, и для адаптивного иммунитета . Некоторые аутоиммунные заболевания, такие как системная красная волчанка , поражают преимущественно женщин, а их наступление совпадает с половым созреванием . В то же время мужские гормоны, в частности, тестостерон , обладают иммуносупрессорными свойствами . На иммунную систему влияют и другие гормоны, такие как глюкокортикоиды , пролактин , соматотропин и производные витамина D .
Гормоны из группы глюкокортикоидов являются важнейшими регуляторами иммунной системы, так как повышают экспрессию противовоспалительных белков (таких как , , SLPI и MAPK-фосфатазы ) и понижают выработку провоспалительных белков. Благодаря выраженному противовоспалительному действию глюкокортикоиды используются в терапии аутоиммунных заболеваний , аллергии , сепсиса . Глюкокортикоиды также задействованы в регуляции развития T-клеток .
Когда T-клетка встречается с патогеном, она выставляет наружу . Благодаря ему T-клетка взаимодействует с активной формой витамина D — стероидным гормоном кальцитриолом . Но этим связь T-клеток с витамином D не ограничивается. T-клетки экспрессируют ген , кодируемый которым фермент превращает производное витамина D кальцидиол в кальцитриол. T-клетки могут выполнять свои защитные функции только после связывания с кальцитриолом. Ген CYP27B1 экспрессируется и некоторыми другими клетками, которые также способны образовывать кальцитриол из кальцидиола: дендритными клетками, макрофагами, а также кератиноцитами .
Предполагается, что прогрессирующее снижение уровня гормонов с возрастом может быть связано с ослабленным иммунитетом у пожилых людей . Кроме того, иммунная система также влияет на эндокринную, в частности, на тиреоидные гормоны . Возрастное снижение иммунитета коррелирует с понижающимся уровнем витамина D у людей преклонного возраста .
Сон и отдых
Сон и отдых оказывают влияние на иммунную систему , в частности, недостаток сна угнетает её функционирование . Сложная петля обратных связей, включающая некоторые цитокины, такие как интерлейкин 1 и , образующиеся во время инфекции, задействована в регуляции медленного сна . Благодаря этому во время инфекции цикл сна может меняться, а именно, увеличиваться доля медленноволнового сна .
У людей, страдающих от недостатка сна, может наблюдаться сниженный по сравнению с обычными людьми иммунный ответ и пониженное образование антител в ответ на инфекцию. Кроме того, нарушения циклов чередования светлого и тёмного времени суток нарушают работу белка , который вовлечён в регуляцию не только циркадных ритмов , но и дифференцировки T-клеток. Нарушение суточных ритмов вместе с нарушениями в работе иммунной системы могут вызывать болезни сердца , астму и .
Помимо отрицательного эффекта нехватки сна на работу иммунной системы, сон и циркадные ритмы оказывают сильное регулирующее воздействие и на врождённый, и на приобретённый иммунитет. Во время медленноволнового сна происходит резкий провал в уровне кортизола , адреналина и норадреналина , из-за чего возрастает концентрация лептина , соматотропина и пролактина. Эти гормоны связаны с образованием провоспалительных цитокинов IL-1, , TNFα и INFγ , которые активируют иммунные клетки, способствуют их пролиферации и дифференцировке. Именно во время медленноволнового сна по ходу медленно развивающегося адаптивного иммунного ответа достигает пика численность недифференцированных или слабо дифференцированных и T-клеток памяти . Кроме того, гормоны, активно продуцирующиеся во время медленного сна (лептин, соматотропин и пролактин), поддерживают взаимодействие антигенпрезентирующих клеток и T-клеток, увеличивают пролиферацию T-хелперов и миграцию наивных T-клеток в лимфоузлы. Считается, что эти же гормоны способствуют формированию долговременной иммунологической памяти, активируя ответ T-хелперов .
Во время бодрствования, напротив, пика достигает численность T-киллеров и NK-клеток, а также концентрация противовоспалительных веществ, таких как кортизол и катехоламины . Существуют две гипотезы относительно того, почему во время сна активируются провоспалительные сигналы в иммунной системе. Во-первых, если бы активный воспалительный ответ происходил во время бодрствования, он бы вызывал серьёзные физические и когнитивные нарушения. Во-вторых, протеканию воспаления во сне может способствовать мелатонин . Во время воспаления образуется огромное количество свободных радикалов ( окислительный стресс ), и мелатонин может противостоять их образованию во время сна .
Питание
Переедание связано с такими заболеваниями, как диабет и ожирение , которые влияют на работу иммунной системы. Умеренное недоедание , а также нехватка некоторых микроэлементов и питательных веществ , могут отрицательно сказываться на работе иммунитета . Имеются данные, что на иммунитет положительно влияют продукты, богатые жирными кислотами , а недостаточное снабжение питательными веществами плода во время беременности может нарушить работу иммунной системы на всю оставшуюся жизнь .
Заживление ран и регенерация
Иммунная система, в особенности её врождённая составляющая, играет важнейшую роль в восстановлении тканей после повреждения . Ключевыми игроками этого процесса являются макрофаги и нейтрофилы, однако важное значение имеют также γδ-T клетки, врождённые лимфоидные клетки и регуляторные T-клетки. Важнейшими факторами заживления раны являются пластичность иммунных клеток и баланс между провоспалительными и противовоспалительными сигналами. Иммунная система вовлечена и в процесс регенерации, в частности, у амфибий . Согласно одной из гипотез, виды, имеющие высокую способность к регенерации, менее иммунокомпетенты, чем виды, имеющие низкий регенеративный потенциал .
Нарушения иммунной системы у человека
Нарушения в работе иммунной системы можно подразделить на три категории: иммунодефициты, аутоиммунные заболевания и реакции гиперчувствительности .
Иммунодефицит
Иммунодефицит возникает при недостаточной эффективности работы иммунной системы, когда один или более её компонентов не функционируют. Активность иммунной системы после 50 лет постепенно снижается, этот процесс называют ( англ. immunosenescence ) . В развитых странах основными причинами снижения эффективности работы иммунной системы являются ожирение, алкоголизм и наркомания . В развивающихся странах нарушения в работе иммунной системы чаще всего вызваны недоеданием. При нехватке белков в рационе нарушается работа клеточных механизмов иммунитета, системы комплемента, снижается образование цитокинов и иммуноглобулинов A (IgA) . Некоторые иммунодефициты имеют наследственную природу , например, , при которой фагоциты разрушают клетки патогенов с низкой эффективностью. Иммунодефицит может быть результатом ВИЧ-инфекции или некоторых онкологических заболеваний .
Аутоиммунные заболевания
Аутоиммунные заболевания связаны с повышенной гиперактивностью иммунной системы, которая начинает атаковать белки самого организма. Таким образом, при аутоиммунных заболеваниях нарушается механизм распознавания своего и чужого. В норме T-клетки, способные распознавать собственные белки организма, уничтожаются с помощью специальных механизмов . Механизм аутоиммунного разрушения клеток и тканей не отличается от того, который реализуется в норме при адаптивном иммунном ответе. К числу распространённых аутоиммунных заболеваний относят ревматоидный артрит , инсулинозависимый сахарный диабет , аутоиммунный тиреоидит .
Гиперчувствительность
Под гиперчувствительностью понимают чрезмерный иммунный ответ на какой-либо антиген. Реакции гиперчувствительности подразделяют на несколько типов в зависимости от их длительности и механизмов, лежащих в их основе. Гиперчувствительность I типа включает немедленные анафилактические реакции, часто связанные с аллергией. Реакции этого типа могут как вызывать небольшой дискомфорт, так и приводить к смерти. Основу гиперчувствительности I типа составляют иммуноглобулины E (IgE), которые вызывают дегрануляцию базофилов и тучных клеток. О гиперчувствительности II типа говорят, когда в организме присутствуют антитела, распознающие его собственные белки и помечающие экспрессирующие их клетки к разрушению. Гиперчувствительность II типа также называют зависимой от антител или цитотоксической гиперчувствительностью, её основу составляют иммуноглобулины G (IgG) и M (IgM). Иммунные комплексы, представляющие собой скопления антигенов, белков комплемента, IgG и IgM, запускают реакции гиперчувствительности III типа. Гиперчувствительность IV типа, также известная как отложенная гиперчувствительность, развивается в течение 2—3 дней. Реакции гиперчувствительности IV типа наблюдаются при многих аутоиммунных и инфекционных заболеваниях, а их основу составляют T-клетки, моноциты и макрофаги .
Медицинские манипуляции
Иммуносупрессия
Препараты-иммуносупрессоры (иммунодепрессанты) используются при лечении аутоиммунных заболеваний, избыточных воспалений, а также для предотвращения отторжения трансплантата после пересадки органа . Для контроля эффектов воспалительных процессов активно применяются противовоспалительные препараты . Одними из самых мощных противовоспалительных средств являются глюкокортикоиды , однако они обладают множеством серьёзных побочных эффектов, среди которых центральное ожирение, гипергликемия , остеопороз , поэтому их приём должен строго контролироваться . Небольшие дозы противовоспалительных препаратов часто используются вместе с цитотоксическими препаратами и препаратами-иммуносупрессорами, такими как метотрексат и азатиоприн . Цитотоксические препараты подавляют иммунный ответ, убивая делящиеся клетки, среди которых и активированные T-клетки. Однако под удар попадают и делящиеся клетки других тканей, поэтому цитотоксические препараты имеют массу побочных эффектов . Препараты-иммуносупрессоры, такие как циклоспорин , подавляют ответ T-клеток на внеклеточные стимулы, ингибируя их сигнальные пути .
Вакцинация
Активный иммунитет может быть создан искусственно за счёт вакцинации. Основной принцип вакцинации, или иммунизации, заключается во введении в организм антигена некоторого патогена с целью развить против него специфический иммунитет без перенесения болезни . Избирательный запуск иммунного ответа с помощью антигена, но не патогена целиком, полагается на естественную специфичность иммунной системы. Вакцинация представляет собой один из самых успешных примеров манипуляции иммунной системой, которая обеспечивает защиту от множества инфекционных заболеваний . Большинство вакцин против вирусных инфекций содержат живой ослабленный вирус, а многие вакцины против бактериальных заболеваний основаны на неклеточных компонентах микроорганизмов, например, безвредных компонентах токсинов . Так как многие антигены, использующиеся в неклеточных вакцинах, не обеспечивают адаптивный иммунный ответ достаточной силы, к большинству антибактериальных вакцин добавляют адъюванты , которые активируют антигенпрезентирующие клетки врождённого иммунитета и увеличивают иммуногенность вакцины .
Предсказание иммуногенности
Организм может развивать нейтрализующий иммунный ответ после введения препаратов, представляющих собой крупные пептиды и белки массой более 500 Да , особенно при повторяющемся введении в крупных дозах. Иногда иммуногенным оказывается не сам препарат, а вещество, вместе с которым он вводится, как иногда бывает с противораковым препаратом таксолом . Для оценки иммуногенности белков и пептидов разработаны вычислительные методы, которые особенно важны для разработки препаратов на основе антител и других веществ пептидной природы, а также для изучения эффектов мутаций белков вирусной оболочки на его вирулентность . Самые первые методы оценки иммуногенности были основаны на наблюдении, что в эпитопах доля гидрофильных аминокислотных остатков гораздо больше, чем гидрофобных . Последние подходы основаны на использовании машинного обучения и баз данных известных эпитопов (как правило, хорошо изученных вирусных белков) в качестве . Область, занимающаяся изучением иммуногенности с помощью биоинформатических подходов, получила название .
Иммунологическая толерантность
Под иммунологической толерантностью понимают отсутствие иммунного ответа на специфический антиген. Перечень антигенов, к которым может развиваться толерантность, практически неотличим от набора антигенов, против которых развивается специфический иммунный ответ. Специфическая неотвечаемость на определённые антигены необходима на некоторых стадиях онтогенеза , а также для нормального протекания беременности у млекопитающих ( иммунологическая толерантность при беременности ). Развитие толерантности к некоторым антигенам происходит на ранних этапах развития организма. Кроме того, толерантность можно искусственно индуцировать на ранних этапах онтогенеза, в частности, за счёт введения антигена в новорождённый организм с не до конца сформированной иммунной системой. Клеточные и молекулярные механизмы толерантности, сформированной в начале развития организма и во взрослом возрасте, нередко отличаются. Так, у новорождённых мышей макрофаги малочисленны, поэтому иммунная система не может развить эффективный ответ. В развитии иммунологической толерантности важную роль играют регуляторные T-клетки , подавляющие T-хелперы .
Иммунология опухолей
Помимо борьбы с патогенами, важная роль иммунной системы заключается в выявлении и уничтожении злокачественных клеток. Клетки опухолей, претерпевшие злокачественную трансформацию, часто экспрессируют поверхностные антигены, отсутствующие у нормальных клеток. Иммунная система воспринимает эти антигены как чужеродные, поэтому они запускают иммунный ответ против злокачественных клеток. Опухолевые антигены могут иметь разное происхождение : они могут происходить от онкогенных вирусов , например, человеческого папилломавируса , вызывающего рак шейки матки и других органов , а другие опухолевые антигены являются, по сути, собственными белками организма, которые в норме присутствуют в клетках на низком уровне, а в опухолевых клетках — на существенно более высоком. Примером может служить фермент тирозиназа , необходимый для синтеза меланина. Высокая экспрессия этого белка приводит к превращению меланоцитов в злокачественные клетки . Третий источник опухолевых антигенов — это белки, в норме регулирующие рост и выживаемость клеток, которые часто мутируют и становятся онкогенами .
Противоопухолевый иммунный ответ заключается преимущественно в уничтожении опухолевых клеток T-киллерами, иногда при участии T-хелперов . Опухолевые антигены презентируются в комплексе с MHC-I подобно вирусным белкам, благодаря чему T-киллеры могут распознать злокачественные клетки . NK-клетки тоже уничтожают опухолевые клетки, особенно если они экспрессируют MHC-I на низком уровне по сравнению с нормой, что происходит с опухолевыми клетками довольно часто . В некоторых случаях в организме образуются антитела к опухолевым антигенам, и злокачественные клетки уничтожаются системой комплемента .
Некоторым опухолям удаётся избежать иммунного ответа . Их клетки экспрессируют MHC-I на низком уровне, благодаря чему их не распознают T-киллеры . Некоторые опухолевые клетки выделяют вещества, подавляющие иммунный ответ, например, цитокин TGFβ , подавляющий активность макрофагов и лимфоцитов . Кроме того, к опухолевым антигенам может развиться иммунологическая толерантность, из-за чего иммунная система утрачивает способность распознавать злокачественные клетки .
Макрофаги могут способствовать росту опухоли из-за цитокинов, которые они используют для привлечения других макрофагов, например, TNFα . Кроме того, сочетание гипоксии , в условиях которой чаще всего находится опухоль , и цитокинов, выделяемых макрофагами, может активизировать метастазирование опухоли .
Эволюция
Многокомпонентная адаптивная иммунная система в ходе эволюции появилась, вероятнее всего, у челюстноротых позвоночных, поскольку у беспозвоночных нет ни лимфоцитов, ни антител . Тем не менее, у многих организмов имеются защитные системы, по принципу работы похожие на адаптивный иммунитет позвоночных. Функциональные аналоги иммунной системы есть даже у бактерий и архей , например, система рестрикции-модификации или система CRISPR /Cas, предназначенные для защиты от вирусов . Систему CRISPR/Cas можно также рассматривать как форму приобретённого иммунитета, поскольку она подразумевает вставку в геном бактерии или археи последовательностей вирусов, с которыми она встречалась, благодаря чему при повтором заражении тем же вирусом быстро возникает специфичная защитная реакция. В системах CRISPR/Cas репликация вирусов подавляется за счёт комплементарного взаимодействия коротких РНК , считываемых со спейсеров в локусе CRISPR, с чужеродными нуклеиновыми кислотами и их последующего разрушения белками Cas . У прокариот есть и другие механизмы противовирусной защиты . Некоторые элементы иммунной системы есть и у одноклеточных эукариот , однако они плохо изучены .
Некоторые компоненты врождённого иммунитета, такие как рецепторы распознавания паттернов, есть практически у всех организмов. Антимикробные пептиды есть у растений и животных, причём у беспозвоночных именно антимикробные пептиды являются главной формой системного иммунитета . У большинства беспозвоночных животных также есть система комплемента и фагоциты. Главную роль в противовирусной защите от вирусов играют ферменты рибонуклеазы и механизм РНК-интерференции, которые консервативны практически у всех эукариот .
В отличие от животных, растения не имеют клеток, способных к фагоцитозу, однако растения всё же имеют иммунную систему, работа которой построена на передаче химических сигналов по всему растительному организму . Отдельные клетки растений также могут реагировать на PAMPs . При заражении участка растительной ткани формируется локальная реакция гиперчувствительности, из-за которой клетки, располагающиеся в зоне поражения, претерпевают апоптоз и становятся неспособны передавать патогены другим клеткам. Организм растения может приобрести резистентность к патогену целиком благодаря защитной реакции, известной как . Важную роль в системном ответе растений на вирусную инфекцию играет РНК-интерференция, блокирующая репликацию вирусов .
Хотя классические молекулы адаптивной иммунной системы, такие как иммуноглобулины и T-клеточные рецепторы, имеются только у челюстноротых, высказываются предположения, что у других организмов есть альтернативные адаптивные иммунные системы. В качестве примера можно привести примитивных бесчелюстных позвоночных, таких как миноги и миксины . У них имеются так называемые ( англ. variable lymphocyte receptors, VLRs ), которые кодируются одним или двумя генами. VLRs связывают антигены похожим на антитела образом и со схожей специфичностью .
Противодействие патогенов
Успех любого патогена зависит от того, насколько успешно он может преодолевать барьеры иммунной системы организма. Поэтому патогены развили несколько механизмов, которые помогают им уклоняться от действия иммунной системы или приводят к её разрушению . Бактерии часто преодолевают защитные барьеры организма, выделяя ферменты, которые их разрушают, например, с помощью систем секреции II типа . Системы секреции III типа обеспечивают связь клетки бактерии с клеткой организма-хозяина посредством полых трубок, благодаря чему белки патогена непосредственно доставляются в клетки организма-хозяина. Часто эти белки предназначены для выключения защитных механизмов хозяина .
Некоторые патогены, например, внутриклеточные, избегают иммунного ответа, «прячась» внутри клеток хозяина. Такие патогены большую часть своего жизненного цикла проводят внутри клеток хозяина, где они защищены от действия иммунных клеток, антител и системы комплемента. К числу внутриклеточных патогенов относятся вирусы, некоторые бактерии (например, Salmonella ) и даже эукариоты ( малярийный плазмодий Plasmodium falciparum и протисты рода Leishmania ). Некоторые бактерии, такие как Mycobacterium tuberculosis , обитают внутри капсул , которые защищают их от лизиса белками комплемента . Многие патогены выделяют вещества, которые сводят на нет иммунный ответ или уводят его в неверном направлении. Некоторые патогенные бактерии формируют биоплёнки , в которых все бактериальные клетки надёжно укрыты от действия иммунной системы. Биоплёнки формируют такие возбудители заболеваний человека, как Pseudomonas aeruginosa и . Другие бактерии синтезируют поверхностные белки, связывающие антитела и инактивирующие их; примером могут служить белок G бактерий рода Streptococcus , белок A Staphylococcus aureus и белок L Peptostreptococcus magnus .
Механизмы избегания адаптивного иммунного ответа устроены более сложно. Простейшим из них является , благодаря которой несущественные части поверхностных эпитопов могут быстро изменяться. Например, у ВИЧ белки вирусной оболочки, необходимые для проникновения в клетку-мишень, постоянно меняются. Частые изменения в антигенах могут объяснить неудачи в создании вакцины против этого вируса . Похожий механизм вариации поверхностных белков использует эукариотический одноклеточный паразит , благодаря чему он может на шаг опережать иммунную систему . Ещё одна часто используемая стратегия заключается в маскировании антигенов. Так, вирионы ВИЧ при отделении от клетки окружают себя липидной оболочкой, происходящей из мембраны клетки-хозяина, благодаря чему иммунной системе оказывается сложнее опознать их как чужеродные объекты .
История изучения

Первые известные записи о существовании иммунитета датируются 430 годом до н. э. , когда в Афинах бушевала эпидемия чумы . Фукидид заметил, что люди, переболевшие чумой во время предыдущей вспышки, могут ухаживать за больными, не заболевая повторно . В X веке персидский врач Абу Бакр Мухаммад ар-Рази впервые ввёл представление об адаптивном иммунитете и описал защитное действие перенесённой оспы против повторного заболевания . Хотя в его представлении иммунитет заключался в выделении «излишков влаги» с кровью во время первого заболевания, благодаря чему предотвращается повторное заболевание, теория ар-Рази смогла объяснить некоторые наблюдения насчёт оспы, имевшиеся на тот момент .
В XVIII веке Пьер Луи де Мопертюи провёл серию экспериментов с ядом скорпиона и обнаружил, что некоторые собаки и мыши были устойчивы к его действию . Дальнейшее развитие представления об иммунитете получили благодаря Луи Пастеру , разработавшему принципы вакцинации и микробную теорию болезней . Теория Пастера противоречила представлениям того времени о заболеваниях, в частности, широко распространённой теории миазмов . Представления Пастера подтвердил в 1891 году Роберт Кох , который в 1905 году получил Нобелевскую премию по физиологии и медицине за установление микробной причины инфекционных болезней . Роль вирусов в заболеваниях человека была показана в 1901 году, когда Уолтер Рид открыл вирус жёлтой лихорадки .
Иммунология бурно развивалась в конце XIX века. В то время интенсивно изучались клеточные и гуморальные основы иммунитета . Особенно важным был вклад Пауля Эрлиха , который предложил для объяснения специфичности взаимодействия антиген-антитело. В конце XIX и начале XX веков происходило противостояние клеточной и гуморальной теорий иммунитета. Клеточную теорию отстаивал Илья Мечников , открывший способность иммунных клеток к фагоцитозу. Сторонники гуморальной теории иммунитета, среди которых были Пауль Эрлих и Эмиль Адольф фон Беринг , утверждали, что компоненты иммунитета являются свободными молекулами, растворёнными в жидкостях организма, а не специализированные клетки. За свои работы в изучении механизмов гуморального иммунитета Эрлих был удостоен Нобелевской премии по физиологии и медицине в 1908 году, которую он разделил с Мечниковым .
В 1950-х годах Фрэнк Макфарлейн Бёрнет сформулировал теорию клональной селекции в иммунной системе . Согласно Бёрнету, запуск иммунного ответа основан на различении «своего» (компонентов здорового организма, не запускающих иммунный ответ) и «чужого» (патогенов и трансплантатов, которые активируют иммунный ответ) . Теория Бёрнета была в дальнейшем преобразована в соответствии с новыми открытиями, касающимися главного комплекса гистосовместимости и его участия в двухшаговой активации T-клеток .
Примечания
- ↑ Beck G. , Habicht G. S. (англ.) // Scientific American. — 1996. — November ( vol. 275 , no. 5 ). — P. 60—63 . — doi : . — .
- O'Byrne K. J. , Dalgleish A. G. (англ.) // British Journal Of Cancer. — 2001. — 17 August ( vol. 85 , no. 4 ). — P. 473—483 . — doi : . — .
- , с. 164.
- Lawson L. J. , Perry V. H. , Gordon S. (англ.) // Neuroscience. — 1992. — Vol. 48 , no. 2 . — P. 405—415 . — doi : . — .
- Ginhoux F. , Lim S. , Hoeffel G. , Low D. , Huber T. (англ.) // Frontiers In Cellular Neuroscience. — 2013. — Vol. 7 . — P. 45—45 . — doi : . — .
- Filiano A. J. , Gadani S. P. , Kipnis J. (англ.) // Brain Research. — 2015. — 18 August ( vol. 1617 ). — P. 18—27 . — doi : . — .
- Gehrmann J. , Matsumoto Y. , Kreutzberg G. W. (англ.) // Brain Research. Brain Research Reviews. — 1995. — March ( vol. 20 , no. 3 ). — P. 269—287 . — doi : . — .
- , с. 14.
- ↑ Litman G. W. , Cannon J. P. , Dishaw L. J. (англ.) // Nature Reviews. Immunology. — 2005. — November ( vol. 5 , no. 11 ). — P. 866—879 . — doi : . — .
- Restifo N. P. , Gattinoni L. (англ.) // Current Opinion In Immunology. — 2013. — October ( vol. 25 , no. 5 ). — P. 556—563 . — doi : . — .
- Kurosaki T. , Kometani K. , Ise W. (англ.) // Nature Reviews. Immunology. — 2015. — March ( vol. 15 , no. 3 ). — P. 149—159 . — doi : . — .
- Smith A. D. (Ed). (англ.) . — Oxford University Press, 2000. — P. . — ISBN 0-19-854768-4 .
- Saji F. , Samejima Y. , Kamiura S. , Koyama M. (англ.) // Reviews Of Reproduction. — 1999. — May ( vol. 4 , no. 2 ). — P. 81—89 . — doi : . — .
- Van de Perre P. (англ.) // Vaccine. — 2003. — 28 July ( vol. 21 , no. 24 ). — P. 3374—3376 . — doi : . — .
- Keller M. A. , Stiehm E. R. (англ.) // Clinical Microbiology Reviews. — 2000. — October ( vol. 13 , no. 4 ). — P. 602—614 . — doi : . — .
- ↑ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walters, Peter. . — Fourth. — New York and London: , 2002. — ISBN 978-0-8153-3218-3 . 18 сентября 2009 года.
- , с. 327—328.
- Boyton R. J. , Openshaw P. J. (англ.) // British Medical Bulletin. — 2002. — Vol. 61 . — P. 1—12 . — doi : . — .
- Agerberth B. , Gudmundsson G. H. (англ.) // Current Topics In Microbiology And Immunology. — 2006. — Vol. 306 . — P. 67—90 . — doi : . — .
- Moreau J. M. , Girgis D. O. , Hume E. B. , Dajcs J. J. , Austin M. S. , O'Callaghan R. J. (англ.) // Investigative Ophthalmology & Visual Science. — 2001. — September ( vol. 42 , no. 10 ). — P. 2347—2354 . — .
- Hankiewicz J. , Swierczek E. (англ.) // Clinica Chimica Acta; International Journal Of Clinical Chemistry. — 1974. — 17 December ( vol. 57 , no. 3 ). — P. 205—209 . — doi : . — .
- Fair W. R. , Couch J. , Wehner N. (англ.) // Urology. — 1976. — February ( vol. 7 , no. 2 ). — P. 169—177 . — doi : . — .
- Yenugu S. , Hamil K. G. , Birse C. E. , Ruben S. M. , French F. S. , Hall S. H. (англ.) // The Biochemical Journal. — 2003. — 1 June ( vol. 372 , no. Pt 2 ). — P. 473—483 . — doi : . — .
- Smith J. L. (англ.) // Journal Of Food Protection. — 2003. — July ( vol. 66 , no. 7 ). — P. 1292—1303 . — doi : . — .
- Gorbach S. L. (англ.) // Annals Of Medicine. — 1990. — February ( vol. 22 , no. 1 ). — P. 37—41 . — .
- Hill L. V. , Embil J. A. (англ.) // CMAJ : Canadian Medical Association Journal = Journal De L'Association Medicale Canadienne. — 1986. — 15 February ( vol. 134 , no. 4 ). — P. 321—331 . — .
- Reid G. , Bruce A. W. (англ.) // Postgraduate Medical Journal. — 2003. — August ( vol. 79 , no. 934 ). — P. 428—432 . — doi : . — .
- Salminen S. J. , Gueimonde M. , Isolauri E. (англ.) // The Journal Of Nutrition. — 2005. — May ( vol. 135 , no. 5 ). — P. 1294—1298 . — doi : . — .
- Reid G. , Jass J. , Sebulsky M. T. , McCormick J. K. (англ.) // Clinical Microbiology Reviews. — 2003. — October ( vol. 16 , no. 4 ). — P. 658—672 . — doi : . — .
- Medzhitov R. (англ.) // Nature. — 2007. — 18 October ( vol. 449 , no. 7164 ). — P. 819—826 . — doi : . — .
- Matzinger P. (англ.) // Science (New York, N.Y.). — 2002. — Vol. 296, no. 5566 . — P. 301—305. — doi : . — .
- Kumar H. , Kawai T. , Akira S. (англ.) // International Reviews Of Immunology. — 2011. — February ( vol. 30 , no. 1 ). — P. 16—34 . — doi : . — .
- Schroder K. , Tschopp J. (англ.) // Cell. — 2010. — 19 March ( vol. 140 , no. 6 ). — P. 821—832 . — doi : . — .
- Beutler B. , Jiang Z. , Georgel P. , Crozat K. , Croker B. , Rutschmann S. , Du X. , Hoebe K. (англ.) // Annual Review Of Immunology. — 2006. — Vol. 24 . — P. 353—389 . — doi : . — .
- Botos I. , Segal D. M. , Davies D. R. (англ.) // Structure (London, England : 1993). — 2011. — 13 April ( vol. 19 , no. 4 ). — P. 447—459 . — doi : . — .
- Takeda K. , Kaisho T. , Akira S. (англ.) // Annual Review Of Immunology. — 2003. — Vol. 21 . — P. 335—376 . — doi : . — .
- Thompson M. R. , Kaminski J. J. , Kurt-Jones E. A. , Fitzgerald K. A. (англ.) // Viruses. — 2011. — June ( vol. 3 , no. 6 ). — P. 920—940 . — doi : . — .
- Platnich J. M. , Muruve D. A. (англ.) // Archives Of Biochemistry And Biophysics. — 2019. — 30 July ( vol. 670 ). — P. 4—14 . — doi : . — .
- ↑ , p. 37—77.
- Withers D. R. (англ.) // Immunology. — 2016. — October ( vol. 149 , no. 2 ). — P. 123—130 . — doi : . — .
- Ryter A. (англ.) // Comparative Immunology, Microbiology And Infectious Diseases. — 1985. — Vol. 8 , no. 2 . — P. 119—133 . — doi : . — .
- Langermans J. A. , Hazenbos W. L. , van Furth R. (англ.) // Journal Of Immunological Methods. — 1994. — 14 September ( vol. 174 , no. 1-2 ). — P. 185—194 . — doi : . — .
- Salzet M. , Tasiemski A. , Cooper E. (англ.) // Current Pharmaceutical Design. — 2006. — Vol. 12 , no. 24 . — P. 3043—3050 . — doi : . — .
- Zen K. , Parkos C. A. (англ.) // Current opinion in cell biology. — 2003. — Vol. 15, no. 5 . — P. 557—564. — .
- ↑ Stvrtinová, Viera; Jakubovský, Ján; Hulín, Ivan. (англ.) . — Computing Centre, Slovak Academy of Sciences: Academic Electronic Press, 1995. — ISBN 80-967366-1-2 . 11 июля 2001 года.
- Rua R. , McGavern D. B. (англ.) // Journal Of Leukocyte Biology. — 2015. — September ( vol. 98 , no. 3 ). — P. 319—332 . — doi : . — .
- Guermonprez P. , Valladeau J. , Zitvogel L. , Théry C. , Amigorena S. (англ.) // Annual review of immunology. — 2002. — Vol. 20. — P. 621—667. — doi : . — .
- Krishnaswamy G. , Ajitawi O. , Chi D. S. (англ.) // Methods In Molecular Biology (Clifton, N.J.). — 2006. — Vol. 315 . — P. 13—34 . — doi : . — .
- Kariyawasam H. H. , Robinson D. S. (англ.) // Seminars In Respiratory And Critical Care Medicine. — 2006. — April ( vol. 27 , no. 2 ). — P. 117—127 . — doi : . — .
- Spits H. , Cupedo T. (англ.) // Annual Review Of Immunology. — 2012. — Vol. 30 . — P. 647—675 . — doi : . — .
- Gabrielli S. , Ortolani C. , Del Zotto G. , Luchetti F. , Canonico B. , Buccella F. , Artico M. , Papa S. , Zamai L. (англ.) // Journal Of Immunology Research. — 2016. — Vol. 2016 . — P. 1376595—1376595 . — doi : . — .
- Rajalingam R. (англ.) // Methods In Molecular Biology (Clifton, N.J.). — 2012. — Vol. 882 . — P. 391—414 . — doi : . — .
- Kawai T. , Akira S. (англ.) // Nature Immunology. — 2006. — February ( vol. 7 , no. 2 ). — P. 131—137 . — doi : . — .
- Miller S. B. (англ.) // Seminars In Arthritis And Rheumatism. — 2006. — August ( vol. 36 , no. 1 ). — P. 37—49 . — doi : . — .
- Ogawa Y. , Calhoun W. J. (англ.) // The Journal Of Allergy And Clinical Immunology. — 2006. — October ( vol. 118 , no. 4 ). — P. 789—798 . — doi : . — .
- Le Y. , Zhou Y. , Iribarren P. , Wang J. (англ.) // Cellular & Molecular Immunology. — 2004. — April ( vol. 1 , no. 2 ). — P. 95—104 . — .
- Martin P. , Leibovich S. J. (англ.) // Trends In Cell Biology. — 2005. — November ( vol. 15 , no. 11 ). — P. 599—607 . — doi : . — .
- ↑ Rus H. , Cudrici C. , Niculescu F. (англ.) // Immunologic Research. — 2005. — Vol. 33 , no. 2 . — P. 103—112 . — doi : . — .
- Degn S. E. , Thiel S. (англ.) // Scandinavian Journal Of Immunology. — 2013. — August ( vol. 78 , no. 2 ). — P. 181—193 . — doi : . — .
- Liszewski M. K. , Farries T. C. , Lublin D. M. , Rooney I. A. , Atkinson J. P. (англ.) // Advances In Immunology. — 1996. — Vol. 61 . — P. 201—283 . — doi : . — .
- Sim R. B. , Tsiftsoglou S. A. (англ.) // Biochemical Society Transactions. — 2004. — February ( vol. 32 , no. Pt 1 ). — P. 21—27 . — doi : . — .
- Pancer Z. , Cooper M. D. (англ.) // Annual Review Of Immunology. — 2006. — Vol. 24 . — P. 497—518 . — doi : . — .
- Holtmeier W. , Kabelitz D. (англ.) // Chemical Immunology And Allergy. — 2005. — Vol. 86 . — P. 151—183 . — doi : . — .
- , p. 139—173.
- Harty J. T. , Tvinnereim A. R. , White D. W. (англ.) // Annual Review Of Immunology. — 2000. — Vol. 18 . — P. 275—308 . — doi : . — .
- ↑ Radoja S. , Frey A. B. , Vukmanovic S. (англ.) // Critical Reviews In Immunology. — 2006. — Vol. 26 , no. 3 . — P. 265—290 . — doi : . — .
- Abbas A. K. , Murphy K. M. , Sher A. (англ.) // Nature. — 1996. — 31 October ( vol. 383 , no. 6603 ). — P. 787—793 . — doi : . — .
- McHeyzer-Williams L. J. , Malherbe L. P. , McHeyzer-Williams M. G. (англ.) // Current Topics In Microbiology And Immunology. — 2006. — Vol. 311 . — P. 59—83 . — doi : . — .
- Kovacs B. , Maus M. V. , Riley J. L. , Derimanov G. S. , Koretzky G. A. , June C. H. , Finkel T. H. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2002. — 12 November ( vol. 99 , no. 23 ). — P. 15006—15011 . — doi : . — .
- Bettelli E. , Carrier Y. , Gao W. , Korn T. , Strom T. B. , Oukka M. , Weiner H. L. , Kuchroo V. K. (англ.) // Nature. — 2006. — 11 May ( vol. 441 , no. 7090 ). — P. 235—238 . — doi : . — .
- Curiel T. J. (англ.) // The Journal Of Clinical Investigation. — 2007. — May ( vol. 117 , no. 5 ). — P. 1167—1174 . — doi : . — .
- Girardi M. (англ.) // The Journal Of Investigative Dermatology. — 2006. — January ( vol. 126 , no. 1 ). — P. 25—31 . — doi : . — .
- Sproul T. W. , Cheng P. C. , Dykstra M. L. , Pierce S. K. (англ.) // International Reviews Of Immunology. — 2000. — Vol. 19 , no. 2-3 . — P. 139—155 . — doi : . — .
- Parker D. C. (англ.) // Annual Review Of Immunology. — 1993. — Vol. 11 . — P. 331—360 . — doi : . — .
- , p. 399—445.
- Wick G. , Hu Y. , Schwarz S. , Kroemer G. (англ.) // Endocrine Reviews. — 1993. — October ( vol. 14 , no. 5 ). — P. 539—563 . — doi : . — .
- Kroemer G. , Brezinschek H. P. , Faessler R. , Schauenstein K. , Wick G. (англ.) // Immunology Today. — 1988. — June ( vol. 9 , no. 6 ). — P. 163—165 . — doi : . — .
- Trakhtenberg E. F. , Goldberg J. L. (англ.) // Science (New York, N.Y.). — 2011. — 7 October ( vol. 334 , no. 6052 ). — P. 47—48 . — doi : . — .
- Veiga-Fernandes H. , Mucida D. (англ.) // Cell. — 2016. — 5 May ( vol. 165 , no. 4 ). — P. 801—811 . — doi : . — .
- (англ.) // Nature Neuroscience. — 2017. — February ( vol. 20 , no. 2 ). — P. 127—127 . — doi : . — .
- Wira, CR; Crane-Godreau M; Grant K. Endocrine regulation of the mucosal immune system in the female reproductive tract // Mucosal Immunology / Ogra P. L., Mestecky J., Lamm M. E., Strober W., McGhee J. R., Bienenstock J.. — San Francisco: Elsevier , 2004. — ISBN 0-12-491543-4 .
- Lang T. J. (англ.) // Clinical Immunology (Orlando, Fla.). — 2004. — December ( vol. 113 , no. 3 ). — P. 224—230 . — doi : . — .
- Moriyama A. , Shimoya K. , Ogata I. , Kimura T. , Nakamura T. , Wada H. , Ohashi K. , Azuma C. , Saji F. , Murata Y. (англ.) // Molecular Human Reproduction. — 1999. — July ( vol. 5 , no. 7 ). — P. 656—661 . — doi : . — .
- Cutolo M. , Sulli A. , Capellino S. , Villaggio B. , Montagna P. , Seriolo B. , Straub R. H. (англ.) // Lupus. — 2004. — Vol. 13 , no. 9 . — P. 635—638 . — doi : . — .
- King A. E. , Critchley H. O. , Kelly R. W. (англ.) // Molecular Human Reproduction. — 2000. — February ( vol. 6 , no. 2 ). — P. 191—196 . — doi : . — .
- Fimmel S. , Zouboulis C. C. (англ.) // The Aging Male : The Official Journal Of The International Society For The Study Of The Aging Male. — 2005. — September ( vol. 8 , no. 3-4 ). — P. 166—174 . — doi : . — .
- Dorshkind K. , Horseman N. D. (англ.) // Endocrine Reviews. — 2000. — June ( vol. 21 , no. 3 ). — P. 292—312 . — doi : . — .
- Nagpal S. , Na S. , Rathnachalam R. (англ.) // Endocrine Reviews. — 2005. — August ( vol. 26 , no. 5 ). — P. 662—687 . — doi : . — .
- Newton R. , Holden N. S. (англ.) // Molecular Pharmacology. — 2007. — October ( vol. 72 , no. 4 ). — P. 799—809 . — doi : . — .
- Rhen T. , Cidlowski J. A. (англ.) // The New England Journal Of Medicine. — 2005. — 20 October ( vol. 353 , no. 16 ). — P. 1711—1723 . — doi : . — .
- Pazirandeh A. , Xue Y. , Prestegaard T. , Jondal M. , Okret S. (англ.) // FASEB Journal : Official Publication Of The Federation Of American Societies For Experimental Biology. — 2002. — May ( vol. 16 , no. 7 ). — P. 727—729 . — doi : . — .
- von Essen M. R. , Kongsbak M. , Schjerling P. , Olgaard K. , Odum N. , Geisler C. (англ.) // Nature Immunology. — 2010. — April ( vol. 11 , no. 4 ). — P. 344—349 . — doi : . — .
- Sigmundsdottir H. , Pan J. , Debes G. F. , Alt C. , Habtezion A. , Soler D. , Butcher E. C. (англ.) // Nature Immunology. — 2007. — March ( vol. 8 , no. 3 ). — P. 285—293 . — doi : . — .
- Hertoghe T. (англ.) // Annals Of The New York Academy Of Sciences. — 2005. — December ( vol. 1057 ). — P. 448—465 . — doi : . — .
- Klein J. R. (англ.) // Experimental Biology And Medicine (Maywood, N.J.). — 2006. — March ( vol. 231 , no. 3 ). — P. 229—236 . — doi : . — .
- Mosekilde L. (англ.) // Clinical Endocrinology. — 2005. — March ( vol. 62 , no. 3 ). — P. 265—281 . — doi : . — .
- Lange T. , Perras B. , Fehm H. L. , Born J. (англ.) // Psychosomatic Medicine. — 2003. — September ( vol. 65 , no. 5 ). — P. 831—835 . — doi : . — .
- Bryant P. A. , Trinder J. , Curtis N. (англ.) // Nature Reviews. Immunology. — 2004. — June ( vol. 4 , no. 6 ). — P. 457—467 . — doi : . — .
- Krueger J. M. , Majde J. A. (англ.) // Annals Of The New York Academy Of Sciences. — 2003. — May ( vol. 992 ). — P. 9—20 . — doi : . — .
- Majde J. A. , Krueger J. M. (англ.) // The Journal Of Allergy And Clinical Immunology. — 2005. — December ( vol. 116 , no. 6 ). — P. 1188—1198 . — doi : . — .
- (8 ноября 2013). Дата обращения: 30 марта 2020. 17 ноября 2013 года.
- ↑ Besedovsky L. , Lange T. , Born J. (англ.) // Pflugers Archiv : European Journal Of Physiology. — 2012. — January ( vol. 463 , no. 1 ). — P. 121—137 . — doi : . — .
- Дата обращения: 28 апреля 2014. Архивировано из 9 мая 2014 года.
- R. M. Suskind, C. L. Lachney, J. N. Udall, Jr. Malnutrition and the Immune Response, in: Dairy products in human health and nutrition , M. Serrano-Ríos, ed. — CRC Press, 1994. — P. 285–300.
- Pond C. M. (англ.) // Prostaglandins, Leukotrienes, And Essential Fatty Acids. — 2005. — July ( vol. 73 , no. 1 ). — P. 17—30 . — doi : . — .
- Langley-Evans S. C. , Carrington L. J. (англ.) // Lupus. — 2006. — Vol. 15 , no. 11 . — P. 746—752 . — doi : . — .
- Park J. E. , Barbul A. (англ.) // American Journal Of Surgery. — 2004. — May ( vol. 187 , no. 5A ). — P. 11—16 . — doi : . — .
- Burzyn D. , Kuswanto W. , Kolodin D. , Shadrach J. L. , Cerletti M. , Jang Y. , Sefik E. , Tan T. G. , Wagers A. J. , Benoist C. , Mathis D. (англ.) // Cell. — 2013. — 5 December ( vol. 155 , no. 6 ). — P. 1282—1295 . — doi : . — .
- Leoni G. , Neumann P. A. , Sumagin R. , Denning T. L. , Nusrat A. (англ.) // Mucosal Immunology. — 2015. — September ( vol. 8 , no. 5 ). — P. 959—968 . — doi : . — .
- Wynn T. A. , Vannella K. M. (англ.) // Immunity. — 2016. — 15 March ( vol. 44 , no. 3 ). — P. 450—462 . — doi : . — .
- Laurent P. , Jolivel V. , Manicki P. , Chiu L. , Contin-Bordes C. , Truchetet M. E. , Pradeu T. (англ.) // Frontiers In Immunology. — 2017. — Vol. 8 . — P. 454—454 . — doi : . — .
- Eming S. A. , Hammerschmidt M. , Krieg T. , Roers A. (англ.) // Seminars In Cell & Developmental Biology. — 2009. — July ( vol. 20 , no. 5 ). — P. 517—527 . — doi : . — .
- Godwin J. W. , Pinto A. R. , Rosenthal N. A. (англ.) // Seminars In Cell & Developmental Biology. — 2017. — January ( vol. 61 ). — P. 71—79 . — doi : . — .
- , с. 364, 375, 380.
- Aw D. , Silva A. B. , Palmer D. B. (англ.) // Immunology. — 2007. — April ( vol. 120 , no. 4 ). — P. 435—446 . — doi : . — .
- ↑ Chandra R. K. (англ.) // The American Journal Of Clinical Nutrition. — 1997. — August ( vol. 66 , no. 2 ). — P. 460—463 . — doi : . — .
- Joos L. , Tamm M. (англ.) // Proceedings Of The American Thoracic Society. — 2005. — Vol. 2 , no. 5 . — P. 445—448 . — doi : . — .
- Copeland K. F. , Heeney J. L. (англ.) // Microbiological Reviews. — 1996. — December ( vol. 60 , no. 4 ). — P. 722—742 . — .
- Miller J. F. (англ.) // Immunologic Research. — 1993. — Vol. 12 , no. 2 . — P. 115—130 . — doi : . — .
- , с. 376—377.
- Ghaffar, Abdul . Microbiology and Immunology On-line . University of South Carolina School of Medicine (2006). Дата обращения: 29 мая 2016. 18 мая 2016 года.
- ↑ , p. 701—748.
- ↑ Taylor A. L. , Watson C. J. , Bradley J. A. (англ.) // Critical Reviews In Oncology/hematology. — 2005. — October ( vol. 56 , no. 1 ). — P. 23—46 . — doi : . — .
- Barnes P. J. (англ.) // European Journal Of Pharmacology. — 2006. — 8 March ( vol. 533 , no. 1-3 ). — P. 2—14 . — doi : . — .
- Masri M. A. (англ.) // Molecular Immunology. — 2003. — July ( vol. 39 , no. 17-18 ). — P. 1073—1077 . — doi : . — .
- . Дата обращения: 30 марта 2020. 2 мая 2008 года.
- Singh M. , O'Hagan D. (англ.) // Nature Biotechnology. — 1999. — November ( vol. 17 , no. 11 ). — P. 1075—1081 . — doi : . — .
- Welling G. W. , Weijer W. J. , van der Zee R. , Welling-Wester S. (англ.) // FEBS Letters. — 1985. — 2 September ( vol. 188 , no. 2 ). — P. 215—218 . — doi : . — .
- Söllner J. , Mayer B. (англ.) // : JMR. — 2006. — May ( vol. 19 , no. 3 ). — P. 200—208 . — doi : . — .
- Flower D. R. , Doytchinova I. A. (англ.) // Applied Bioinformatics. — 2002. — Vol. 1 , no. 4 . — P. 167—176 . — .
- , с. 316—321.
- ↑ Andersen M. H. , Schrama D. , Thor Straten P. , Becker J. C. (англ.) // The Journal Of Investigative Dermatology. — 2006. — January ( vol. 126 , no. 1 ). — P. 32—41 . — doi : . — .
- Boon T. , van der Bruggen P. (англ.) // The Journal Of Experimental Medicine. — 1996. — 1 March ( vol. 183 , no. 3 ). — P. 725—729 . — doi : . — .
- Ljubojevic S. , Skerlev M. (англ.) // Clinics In Dermatology. — 2014. — March ( vol. 32 , no. 2 ). — P. 227—234 . — doi : . — .
- Castelli C. , Rivoltini L. , Andreola G. , Carrabba M. , Renkvist N. , Parmiani G. (англ.) // Journal Of Cellular Physiology. — 2000. — March ( vol. 182 , no. 3 ). — P. 323—331 . — doi : . — .
- ↑ Romero P. , Cerottini J. C. , Speiser D. E. (англ.) // Advances In Immunology. — 2006. — Vol. 92 . — P. 187—224 . — doi : . — .
- ↑ Guevara-Patiño J. A. , Turk M. J. , Wolchok J. D. , Houghton A. N. (англ.) // Advances In Cancer Research. — 2003. — Vol. 90 . — P. 157—177 . — doi : . — .
- Renkvist N. , Castelli C. , Robbins P. F. , Parmiani G. (англ.) // Cancer Immunology, Immunotherapy : CII. — 2001. — March ( vol. 50 , no. 1 ). — P. 3—15 . — doi : . — .
- Gerloni M. , Zanetti M. (англ.) // Springer Seminars In Immunopathology. — 2005. — June ( vol. 27 , no. 1 ). — P. 37—48 . — doi : . — .
- ↑ Seliger B. , Ritz U. , Ferrone S. (англ.) // International Journal Of Cancer. — 2006. — 1 January ( vol. 118 , no. 1 ). — P. 129—138 . — doi : . — .
- Hayakawa Y. , Smyth M. J. (англ.) // Advances In Cancer Research. — 2006. — Vol. 95 . — P. 293—322 . — doi : . — .
- ↑ Syn N. L. , Teng MWL , Mok TSK , Soo R. A. (англ.) // The Lancet. Oncology. — 2017. — December ( vol. 18 , no. 12 ). — P. e731—741 . — doi : . — .
- ↑ Seliger B. (англ.) // BioDrugs : Clinical Immunotherapeutics, Biopharmaceuticals And Gene Therapy. — 2005. — Vol. 19 , no. 6 . — P. 347—354 . — doi : . — .
- Frumento G. , Piazza T. , Di Carlo E. , Ferrini S. (англ.) // Endocrine, Metabolic & Immune Disorders Drug Targets. — 2006. — September ( vol. 6 , no. 3 ). — P. 233—237 . — doi : . — .
- Stix G. (англ.) // Scientific American. — 2007. — July ( vol. 297 , no. 1 ). — P. 60—67 . — doi : . — .
- Bickle T. A. , Krüger D. H. (англ.) // Microbiological Reviews. — 1993. — June ( vol. 57 , no. 2 ). — P. 434—450 . — .
- Barrangou R., Fremaux C., Deveau H., Richards M., Boyaval P., Moineau S., Romero D. A., Horvath P. // Science. — 2007. — Vol. 315, no. 5819. — P. 1709—1712. — doi : . — .
- Brouns S. J. , Jore M. M. , Lundgren M. , Westra E. R. , Slijkhuis R. J. , Snijders A. P. , Dickman M. J. , Makarova K. S. , Koonin E. V. , van der Oost J. (англ.) // Science (New York, N.Y.). — 2008. — 15 August ( vol. 321 , no. 5891 ). — P. 960—964 . — doi : . — .
- Hille F. , Charpentier E. (англ.) // Philosophical Transactions Of The Royal Society Of London. Series B, Biological Sciences. — 2016. — 5 November ( vol. 371 , no. 1707 ). — doi : . — .
- Koonin E. V. (англ.) // Biology Direct. — 2017. — 10 February ( vol. 12 , no. 1 ). — P. 5—5 . — doi : . — .
- Bayne C. J. (англ.) // Integrative And Comparative Biology. — 2003. — April ( vol. 43 , no. 2 ). — P. 293—299 . — doi : . — .
- Stram Y. , Kuzntzova L. (англ.) // Virus Genes. — 2006. — June ( vol. 32 , no. 3 ). — P. 299—306 . — doi : . — .
- ↑ Schneider, David . Stanford University Department of Microbiology and Immunology. Дата обращения: 1 января 2007. 6 января 2017 года.
- Jones J. D. , Dangl J. L. (англ.) // Nature. — 2006. — 16 November ( vol. 444 , no. 7117 ). — P. 323—329 . — doi : . — .
- Baulcombe D. (англ.) // Nature. — 2004. — 16 September ( vol. 431 , no. 7006 ). — P. 356—363 . — doi : . — .
- Alder M. N. , Rogozin I. B. , Iyer L. M. , Glazko G. V. , Cooper M. D. , Pancer Z. (англ.) // Science (New York, N.Y.). — 2005. — 23 December ( vol. 310 , no. 5756 ). — P. 1970—1973 . — doi : . — .
- Finlay B. B. , McFadden G. (англ.) // Cell. — 2006. — 24 February ( vol. 124 , no. 4 ). — P. 767—782 . — doi : . — .
- Cianciotto N. P. (англ.) // Trends In Microbiology. — 2005. — December ( vol. 13 , no. 12 ). — P. 581—588 . — doi : . — .
- Winstanley C. , Hart C. A. (англ.) // Journal Of Medical Microbiology. — 2001. — February ( vol. 50 , no. 2 ). — P. 116—126 . — doi : . — .
- Finlay B. B. , Falkow S. (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 1997. — June ( vol. 61 , no. 2 ). — P. 136—169 . — .
- Kobayashi H. (англ.) // Treatments In Respiratory Medicine. — 2005. — Vol. 4 , no. 4 . — P. 241—253 . — doi : . — .
- Housden N. G. , Harrison S. , Roberts S. E. , Beckingham J. A. , Graille M. , Stura E. , Gore M. G. (англ.) // Biochemical Society Transactions. — 2003. — June ( vol. 31 , no. Pt 3 ). — P. 716—718 . — doi : . — .
- Burton D. R. , Stanfield R. L. , Wilson I. A. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2005. — 18 October ( vol. 102 , no. 42 ). — P. 14943—14948 . — doi : . — .
- Taylor J. E. , Rudenko G. (англ.) // Trends In Genetics : TIG. — 2006. — November ( vol. 22 , no. 11 ). — P. 614—620 . — doi : . — .
- Cantin R. , Méthot S. , Tremblay M. J. (англ.) // Journal Of Virology. — 2005. — June ( vol. 79 , no. 11 ). — P. 6577—6587 . — doi : . — .
- Retief F. P. , Cilliers L. (англ.) // South African Medical Journal = Suid-Afrikaanse Tydskrif Vir Geneeskunde. — 1998. — January ( vol. 88 , no. 1 ). — P. 50—53 . — .
- Doherty M. , Robertson M. J. (англ.) // Trends In Immunology. — 2004. — December ( vol. 25 , no. 12 ). — P. 623—631 . — doi : . — .
- . — Academic Press , 1989. — С. —7. — ISBN 978-0-08-092583-7 .
- Ostoya P. (фр.) // Revue d'histoire des sciences et de leurs applications. — 1954. — Vol. 7 , n o 1 . — P. 60—78 . — ISSN . — doi : .
- Plotkin S. A. (англ.) // Nature Medicine. — 2005. — April ( vol. 11 , no. 4 Suppl ). — P. 5—11 . — doi : . — .
- . Дата обращения: 30 марта 2020. 10 декабря 2006 года.
- . Дата обращения: 30 марта 2020. Архивировано из 23 октября 2007 года.
- ; Translated by F.G. Binnie. . — Cambridge University Press , 1905. . — «history of humoral immunity.».
- . Дата обращения: 30 марта 2020. 19 февраля 2007 года.
- Burnet F. M. The Clonal Selection Theory of Acquired Immunity (англ.) . — Cambridge: Cambridge University Press , 1959.
- Burnet F. M. Cellular Immunology: Self and Notself. — Cambridge: Cambridge University Press , 1969.
- Bretscher P. , Cohn M. (англ.) // Science (New York, N.Y.). — 1970. — 11 September ( vol. 169 , no. 3950 ). — P. 1042—1049 . — doi : . — .
Литература
- Бурместер Г.-Р., Пецутто А. Наглядная иммунология. — М. : БИНОМ. Лаборатория знаний, 2014. — 320 с. — ISBN 978-5-9963-1731-8 .
- Галактионов В. Г. Иммунология. — М. : Издат. центр «Академия», 2004. — 528 с. — ISBN 5-7695-1260-1 .
- Kenneth Murphy, Casey Weaver. Janeway's Immunobiology. — Garland Science, 2017. — ISBN 978-0-8153-4505-3 .
Ссылки
- от 2 мая 2019 на Wayback Machine from the University of Hartford (англ.)
- from the University of South Carolina (англ.)
- : Какие агенты внутри нас защищают организм от угроз : гид / Научн. консульт. Николай Никитин (докт. мед. наук, проф. каф. вирусологии биофака МГУ). — Постнаука. — 2023.
|
|
|
|---|---|
| Словари и энциклопедии |
|
| В библиографических каталогах |
|
|
|||||
|---|---|---|---|---|---|
| Органы | |||||
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
- 2020-10-30
- 1









