Interested Article - Нейтрофилы

- 2020-01-19
- 1
Нейтрофи́лы , или нейтрофи́льные гранулоци́ты , или нейтрофи́льные сегментоя́дерные гранулоци́ты , или полиморфонуклеа́рные нейтрофи́лы , — наиболее многочисленная группа гранулоцитов , на долю которой приходится от 40 % до 70 % всех лейкоцитов у человека . Нейтрофилы являются частью врождённого иммунитета , их основная функция — фагоцитоз патогенных микроорганизмов ( бактерий , грибков , простейших ) и продуктов распада тканей организма .
Нейтрофилы представляют собой очень подвижные клетки, которые проникают даже в те ткани, которые недоступны для других лейкоцитов. В зависимости от морфологии ядра нейтрофилы подразделяют на палочкоядерные (незрелые) и сегментоядерные (зрелые) нейтрофилы . Развитие нейтрофилов находится под контролем цитокинов , прежде всего , а также GM-CSF , IL-3 и IL-6 . В условиях воспалительного ответа количество нейтрофилов увеличивается под действием и .
Нейтрофилы являются основными фагоцитами кровотока , но в ходе острого воспаления интенсивно мигрируют в очаг воспаления . Они проникают через стенки кровеносных сосудов и движутся по градиенту различных провоспалительных молекул в ходе хемотаксиса . Нейтрофилы — самые многочисленные клетки, входящие в состав гноя , именно они придают ему беловатый или желтоватый цвет .
Структура
Нейтрофилы — наиболее многочисленный подтип лейкоцитов у человека; каждый день в организме человека образуется порядка 10 11 новых нейтрофилов. В норме количество нейтрофилов в литре крови составляет от 2—2,5 до 7,5 миллиардов клеток. У людей африканского и средневосточного происхождения количество нейтрофилов может быть меньше 2,5 миллиардов на литр . В кровотоке находится лишь 1—2 % зрелых нейтрофилов, все остальные локализованы в тканях. Кроме того, в крови здорового человека присутствуют незрелые палочкоядерные нейтрофилы в количестве 0,04—0,3 × 10 9 на литр, что соответствует 1—6 % всех нейтрофилов . У человека повышение содержания палочкоядерных нейтрофилов в крови свидетельствует об остром воспалении, лейкозе или повреждении костного мозга , однако у некоторых животных существенная доля незрелых нейтрофилов в крови является нормой .
В мазках крови нейтрофилы имеют диаметр от 12 до 15 мкм . В суспензии человеческие нейтрофилы достигают 7—9 мкм в диаметре . Неактивированные нейтрофилы, плавающие в кровотоке, имеют сферическую форму; после активации форма нейтрофилов меняется, становится амёбоидной , появляются псевдоподии , с помощью которых нейтрофилы могут захватывать антигены .
На поверхности нейтрофилов присутствуют молекулы CD13 , служащей рецептором для некоторых вирусов , CD14 (рецептор липополисахарида ), β 2 - интегрины ( LFA-1 , Mac-1 и p155/95), Fc-рецепторы ( и CD16 ), ( , Mac-1, ) и рецепторы факторов хемотаксиса. Нейтрофилы постоянно экспрессируют главный комплекс гистосовместимости I класса ( ), а под действием некоторых цитокинов, таких как GM-CSF, начинают экспрессировать . Нейтрофилы несут рецепторы для ключевых факторов, влияющих на их развитие, миграцию и активацию: G-CSF (главный регулятор развития нейтрофилов), IL-17 и IL-23, основного фактора хемотаксиса IL-8 (рецепторы CXCR1 и CXCR2 ), а также хемокина SDF-1 , определяющего связь нейтрофилов с тканями (соответствующий ему рецептор известен как CXCR4 ) .
У человека выделяют пять групп антигенов нейтрофилов, которые обозначают HNA 1—5. Три антигена группы HNA-1 (a, b, c) располагаются на Fc-рецепторе CD16. Единственный антиген группы HNA-2, HNA-2a, входит в состав поверхностного гликопротеина CD177 . Антигены группы HNA-3, a и b, входят в состав белка . Группы HNA-4 и HNA-5 содержат по два антигена, a и b, которые располагаются на интегрине β 2 .
Морфология ядра нейтрофилов


Подобно базофилам и эозинофилам , зрелые нейтрофилы имеют дольчатое клеточное ядро из 2—5 сегментов, соединённых тонкими перемычками. У некоторых животных ядро зрелых нейтрофилов не имеет отчётливой сегментации . По мере созревания нейтрофила ядрышко исчезает (исчезновение ядрышка происходит лишь у некоторых типов дифференцированных клеток, имеющих ядро). Центр ядра составляет эухроматин , а гетерохроматин сосредоточен на периферии. У женщин некоторые нейтрофилы несут инактивированную X-хромосому в виде тельца Барра , которое выглядит как придаток в виде барабанной палочки у одного из сегментов ядра . Поскольку промоторы многих генов в ядре нейтрофила недоступны для дифференцировочных факторов, экспрессия генов и возможность к биосинтезу макромолекул у нейтрофилов ограничены. Тем не менее нейтрофилы всё же сохраняют способность к биосинтезу, хотя и ограниченному. У незрелых палочкоядерных нейтрофилов ядро не подразделено на сегменты, хотя содержит уплотнённый хроматин .
Аппарат Гольджи , как правило, слабо развит, митохондрии и рибосомы редки, шероховатый эндоплазматический ретикулум (ЭПР) отсутствует. В цитоплазме имеется от 50 до 200 гранул, из которых приблизительно 10—20 % (по некоторым оценкам, до трети) составляют азурофильные гранулы , оставшиеся 80—90 % приходится на специфические гранулы. Азурофильные гранулы окружены одинарной мембраной , они более крупные, чем специфические, и имеют диаметр около 0,4 мкм, окрашиваются в фиолетово-красный цвет. Они представляют собой первичные лизосомы , содержат электронно -плотную сердцевину и ферменты , активные при низких значениях pH : кислую фосфатазу , , миелопероксидазу , которая продуцирует из перекиси водорода молекулярный кислород , имеющий антибактериальные свойства, , , , β-галактозидазу , , , лизоцим , нейтральные протеазы ( катепсин G , эластаза , , ). Кроме того, азурофильные гранулы содержат антимикробные пептиды из числа дефензинов , , гранулофизин, лактоферрин , кислые гликозаминогликаны . Поскольку в ходе дифференцировки нейтрофилов в костном мозге азурофильные гранулы появляются раньше специфических, их также называют первичными гранулами . азурофильных гранул — миелопероксидаза и мембранная молекула CD63 .
, или вторичные гранулы — более мелкие (диаметр около 20 мкм), светлые, многочисленные. Они электронно-прозрачны, но иногда содержат кристаллоид. В специфических гранулах содержатся белки с и бактерицидными свойствами — лизоцим, щелочную фосфатазу , лактоферрин , а также белок , связывающий витамин B 12 . В специфических гранулах в больших количествах содержится фермент NADPH-оксидаза , которая катализирует образование активных форм кислорода , выступающих в роли главных бактерицидных факторов фагоцитов . Кроме того, в состав специфических гранул входят разнообразные ферменты, разрушающие внеклеточный матрикс , такие как коллагеназы . Молекулярными маркерами специфических гранул являются лактоферрин и мембранная молекула CD66 .
Помимо первичных и вторичных гранул в нейтрофилах присутствуют третичные желатиназные и секреторные гранулы, или везикулы . Желатиназные гранулы содержат , также известную как матриксная металлопротеиназа 9 (MMP9). Они мельче специфических гранул и легче подвергаются экзоцитозу . В мембранах и специфических, и желатиназных гранул имеется флавоцитохром b558, который входит в состав NADPH-оксидазы. Секреторные везикулы содержат щелочную фосфатазу и несут на своей поверхности Fc-рецептор CD16, рецепторы комплемента (в том числе CD35), а также интегрины CD11b / CD18 , CD11c /CD18 и молекулы CD15 и CD14 . После того как в ходе экзоцитоза мембрана секреторной везикулы встроится в клеточную мембрану нейтрофила, перечисленные рецепторы могут быть моментально использованы клеткой , что может резко изменить её фенотип .
В таблице ниже перечислены основные компоненты мембран и матрикса гранул нейтрофилов .
| Компартмент гранулы | Азурофильные (первичные) гранулы | Специфические (вторичные) гранулы | Желатиназные (третичные) гранулы | Секреторные гранулы |
|---|---|---|---|---|
| Мембрана | CD63 , CD68 | CD15, CD66, CD67, , CD120 , b558 | CD11b/CD18, FPR, b558 | CD11b/CD18, CD13, CD14, CD16, FPR, CD35 |
| Матрикс | , катепсин G, эластаза, азуроцидин, лизоцим, BPI, α-дефензины | Лизоцим, кателицидин, BPI, , лактоферрин, фосфолипаза A2 , коллагеназа | Желатиназа B | Азуроцидин, альбумин , щелочная фосфатаза |
Нейтрофилы содержат гликоген , и основную роль в получении энергии у нейтрофилов играет гликолиз . Цикл Кребса и окислительное фосфорилирование вносят меньший вклад в энергообеспечение нейтрофила, о чём свидетельствует небольшое количество митохондрий у данного типа клеток. Способность нейтрофилов выживать в анаэробных условиях очень важна для их функционирования, поскольку позволяет им убивать и фагоцитировать бактерии даже в бедных кислородом тканях, например, в повреждённых или некротизированных тканях .
Развитие

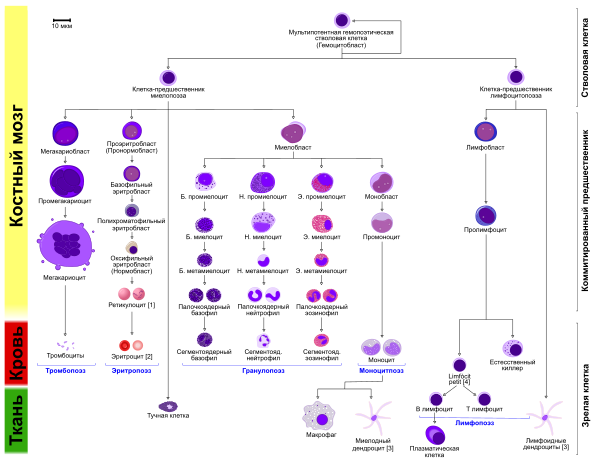
Нейтрофилы образуются в костном мозге из гемопоэтических стволовых клеток . Гемопоэтическая стволовая клетка даёт начало клетке — родоначальнице гранулоцитарного, эритроцитарного , моноцитарного и мегакариоцитарного рядов гемопоэза, которая, в свою очередь, дает начало олигопотентной клетке-предшественнице моноцитов и нейтрофилов. От неё происходит унипотентная клетка-предшественница нейтрофилов, дающая начало миелобластам . Композиция её поверхностных маркеров описывается как CD34 + CD33 +, а также рецепторы для GM-CSF, G-CSF, IL-1, IL-3, IL-6, , IL-12 . Далее последовательность клеток-предшественников выглядит следующим образом: миелобласт → промиелоцит → нейтрофильный миелоцит → метамиелоцит → палочкоядерный нейтрофил → сегментоядерный нейтрофил. Первичные гранулы появляются на стадии промиелоцитов, вторичные — на стадии миелоцитов. До стадии метамиелоцитов предшественники делятся митозом , а метамиелоциты и последующие стадии лишены способности к делению. На стадии метамиелоцитов увеличивается количество специфических гранул в цитоплазме, дальнейшее созревание ядра приводит появлению палочкоядерных нейтрофилов. Сегментация ядра палочкоядерных нейтрофилов приводит к их преобразованию в зрелые сегментоядерные нейтрофилы. Полный период развития нейтрофилов у человека составляет около 14 суток, из которых 7,5 суток приходятся на пролиферацию , а 6,5 — на постмитотическую дифференцировку . У взрослого человека за сутки из костного мозга в кровь выходит от 5 × 10 10 до 10 11 зрелых нейтрофилов .
Срок жизни неактивированного нейтрофила в кровотоке составляет, по разным оценкам, от 5 до 135 часов . При активации нейтрофилы приобретают способность протискиваться через эндотелий сосудов и мигрируют в ткани, где они живут от одного до двух дней. Около 30 % нейтрофилов, покидающих кровоток, мигрируют в костный мозг и печень , 20 % направляются в лёгкие , 15 % — в селезёнку . Главные факторы хемотаксиса нейтрофилов, направляющие их перемещение в ткани, — и IL-8. В процессе миграции нейтрофилов участвуют молекулы адгезии , а именно, β 2 -интегрины, P - и E-селектины , а также секретируемый нейтрофилами фермент . Нейтрофилы гораздо многочисленнее долгоживущих макрофагов , и патоген, проникнувший в организм, прежде всего сталкивается именно с нейтрофилами. Через 3—5 суток пребывания в тканях нейтрофилы подвергаются апоптозу и поглощаются резидентными макрофагами. Некоторые исследователи считают, что короткий срок жизни нейтрофилов является эволюционной адаптацией . Со смертью фагоцита погибают патогены, сохраняющиеся в нём, что защищает организм. Кроме того, ввиду высокой токсичности веществ, выделяемых нейтрофилами для борьбы с инфекцией , по отношению к тканям организма, быстрая гибель нейтрофилов обеспечивает действие противомикробных веществ только в очаге воспаления и защищает остальные ткани организма . Существуют сведения, указывающие на возможность перехода нейтрофилов в тканях в долгоживущую форму и даже в макрофаги .
Функции

Воспалённые или повреждённые участки соединительной ткани требуют немедленной миграции разнообразных лейкоцитов, в том числе нейтрофилов, в очаг повреждения для удаления патогенных микроорганизмов и восстановления ткани. Наиболее хорошо процесс миграции в ткани изучен для нейтрофилов, которые прибывают в очаг воспаления первыми, существенно быстрее моноцитов, и способны развивать защитные метаболические реакции (в частности, « », сопровождающийся продукцией активных форм кислорода) в течение секунд. Активация нейтрофилов сопровождается в первую очередь высвобождением содержимого секреторных гранул. При развитии локального воспаления макрофаги, активированные бактериями или повреждениями ткани, выделяют , такие как IL-1 или фактор некроза опухоли α (TNF-α). Также к факторам хемотаксиса нейтрофилов относят компоненты комплемента , вещества, выделяемые тучными клетками , , эндотоксины и бактериальные пептиды , а также наполненные лизосомы, выходящие в ткань при распаде погибших нейтрофилов и макрофагов . Под действием провоспалительных цитокинов в эндотелиальных клетках, выстилающих ближайшие к очагу воспаления посткапиллярные венулы , увеличивается количество селектинов на поверхности, обращённой в просвет сосуда. Нейтрофилы, циркулирующие в посткапиллярных венулах и имеющие подходящий набор поверхностных гликопротеинов, связываются с селектинами на поверхности эндотелиальных клеток. На данном этапе связывание с эндотелием непрочное, и нейтрофилы продолжают «катиться» по поверхности эндотелия. Провоспалительные цитокины запускают экспрессию интегринов в катящихся нейтрофилах и лигандов интегринов, известных как ICAM-1 , на поверхностях эндотелиальных клеток. В то же время межклеточные контакты между клетками эндотелия ослабляются, и эндотелий становится более проницаемым. За счёт интегринов нейтрофилы прочно связываются с эндотелием и прекращают качение, и в конечном счёте приступают к прохождению через эндотелий с помощью псевдоподий (этот процесс известен как диапедез ). Проникновение нейтрофилов через эндотелий облегчается благодаря длинному и сегментированному ядру. Прохождение нейтрофилов через эндотелий и дальнейшая миграция к очагу воспаления стимулируются хемокинами . Базальные мембраны нейтрофилы преодолевают благодаря выделению содержимого желатиназных гранул .
Координированное перемещение нейтрофилов в очаг острого воспаления называют . Масштаб роения и его длительность определяются многими факторами, среди которых размер повреждённой области ткани и наличие патогенов . Явление роения нейтрофилов было изучено преимущественно у мышей на ткани уха и у рыбок данио-рерио .

Активированные нейтрофилы при встрече с микроорганизмами поглощают их с помощью псевдоподий, и микроорганизм оказывается внутри фагосомы . В течение нескольких секунд после активации нейтрофила изменяется его мембранный потенциал , в клетку входят ионы натрия и кальция , изменяется . Примерно через 30 секунд после поглощения частицы специфические гранулы сливаются с фагосомой, изливая в неё своё содержимое, после чего фагосома дополнительно закисляется с помощью мембранных протонных помп . С закисленными фагосомами сливаются азурофильные гранулы (примерно через 1—3 минуты после поглощения частицы). В ходе фагоцитоза в нейтрофиле образуются активные формы кислорода, такие как супероксид - анион и перекись водорода, и другие компоненты гранул с бактерицидными свойствами. Активные формы кислорода используются для уничтожения бактериальной клетки, наряду с содержимым специфических и азурофильных гранул. Белок лактоферрин, входящий в состав специфических гранул, не убивает бактерию напрямую, а прочно связывает ионы железа , делая их недоступными для бактерии и, в результате, приводя к её смерти. Содержимое гранул (в частности, азурофильных) может высвобождаться в ходе , после которой восстановления гранул не происходит. Комбинация описанных механизмов способна убить практически любую бактерию, и погибшие бактериальные клетки впоследствии расщепляются лизосомными ферментами. Наиболее чувствительны к компонентам гранул нейтрофилов дрожжи ( Candida ) и бактерии из числа стрептококков и стафилококков . Нейтрофилы, погибшие апоптозом, бактерии, полуразрушенные останки клеток и тканевая жидкость формируют густой гной белого или жёлтого цвета .

Ещё одна форма противомикробной активности нейтрофилов заключается в особом типе программируемой клеточной гибели , характерном для нейтрофилов, — нетозе . При нетозе погибающий нейтрофил выбрасывает свою ДНК наружу в виде так называемых внеклеточных ловушек нейтрофилов (от англ. neutrophil extracellular traps, NETs ). Внеклеточные ловушки нейтрофилов состоят из хроматина и сериновых протеаз и способны иммобилизовать и убивать микробные клетки . Таким образом, внеклеточные ловушки нейтрофилов также противодействуют распространению клеток патогенов по тканям. При сепсисе массовый нетоз происходит непосредственно в кровеносных сосудах . Образование внеклеточных ловушек нейтрофилов может вносить вклад в развитие ряда воспалительных заболеваний, таких как преэклампсия , а их образование в кровеносных сосудов может приводить к формированию тромбов , в том числе в коронарных артериях . В 2018 году было показано, что один и тот же нейтрофил может подвергаться и апоптозу, и нетозу одновременно; этот вид программируемой клеточной гибели нейтрофилов получил название апонетоз .
Многие исследователи выделяют две функционально различные субпопуляции нейтрофилов на основании различных интенсивности образования активных форм кислорода, проницаемости мембраны, активности ферментов гранул и способности к инактивации. Нейтрофилы с повышенной проницаемостью мембран называют нейтрофилами-киллерами. Нейтрофилы-киллеры интенсивно генерируют активные формы кислорода и инактивируются после взаимодействия с субстратом. Нейтрофилы второй субпопуляции образуют активные формы кислорода менее интенсивно, не прикрепляются к субстрату и не инактивируются .
Активность нейтрофилов зависит от возраста организма человека. У новорождённого нет адекватной продукции нейтрофилов, а в старческом возрасте способность нейтрофилов к фагоцитозу ограничена .
Нейтрофилы экспрессируют и продуцируют широкий спектр цитокинов, среди которых хемокины, , провоспалительные цитокины ( IL-1α , IL-1β , IL-6, IL-7, IL-18 , и другие), иммунорегуляторные цитокины (IL-12, , , , и другие), противовспалительные цитокины ( IL-1ra , TGFβ1 , ), факторы ангиогенеза и фиброгенеза ( VEGF , BV8, , , , , ангиопоэтин ), цитокины суперсемейства фактора некроза опухоли (TNF) и некоторые другие цитокины, такие как PBEF , , мидкин , , , эндотелин . За счёт выделения разнообразных цитокинов нейтрофилы могут быть вовлечены в процессы, не связанные с иммунной защитой, такие как гемопоэз, ангиогенез и заживление ран. Кроме того, нейтрофилы могут участвовать в развитии некоторых аутоиммунных и злокачественных заболеваний .
Иммуносупрессорные популяции нейтрофилов задействованы в поддержании иммунологической толерантности при беременности . В частности, иммуносупрессорные нейтрофилы низкой плотности могут подавлять пролиферацию CD4 + и CD8 + T-лимфоцитов , подвергая их воздействию активных форм кислорода. Кроме того, они подавляют цитотоксическую активность естественных киллеров и экспрессию Toll-подобных рецепторов на моноцитах . Нейтрофилы, располагающиеся в во время второго триместра беременности , стимулируют в ней дополнительный ангиогенез .
Клиническое значение

Состояние, при котором количество нейтрофилов снижено, называется . Нейтропения может быть наследственной или приобретённой, как при некоторых видах апластической анемии или лейкозов . Нейтропения может развиться вследствие приёма определённых препаратов, в частности, противораковой химиотерапии . Пациенты с нейтропенией отличаются повышенной уязвимостью к инфекционным заболеваниям бактериальной и грибковой природы. Чаще всего пациенты с нейтропенией страдают от таких заболеваний, как воспаление подкожной жировой клетчатки , фурункулёз , пневмония и септицемия . Степень нейтропении определяют с помощью параметра, известного как ( англ. Absolute neutrophil count, ANC ). Состояние с ANC < 1500 клеток / мм³ считается нейтропенией, а с ANC < 500 клеток / мм³ — тяжёлой нейтропенией .

Состояние, противоположное нейтропении, при котором количество нейтрофилов в крови повышено, называют нейтрофилией . Наиболее частой причиной нейтрофилии являются бактериальные инфекции, особенно сопровождающиеся гнойным воспалением . Повышение уровня нейтрофилов происходит при любом остром воспалении, поэтому нейтрофилия может появиться в результате инфаркта миокарда или ожога , а также аппендицита и спленэктомии . Уровень нейтрофилов может повышаться при некоторых злокачественных заболеваниях крови , таких как хронический миелоидный лейкоз , при котором происходит неконтролируемая пролиферация лейкоцитов .
Нейтрофилы, будучи иммунными клетками, принимают участие во взаимодействии с клетками опухолей . Нейтрофилы, инфильтрованные в опухоль, имеют повышенную, по сравнению с нейтрофилами в обычных тканях, продолжительность жизни. Нейтрофилы могут играть ведущую роль в онкогенезе за счёт взаимодействия с другими иммунными клетками , реагирующими на появление и рост опухоли. Нейтрофилы могут стимулировать пролиферацию опухолевых клеток и ангиогенез в опухоли, активировать метастазирование и подавлять иммунный ответ на злокачественные клетки. В то же время нейтрофилы могут обладать и противоопухолевой активностью .
Известно несколько наследственных расстройств, связанных с дисфункцией нейтрофилов. В некоторых случаях подвижность нейтрофилов снижена за счёт нарушений в полимеризации актина , а при недостаточной экспрессии гена, кодирующего NADPH-оксидазу, нейтрофилы теряют способность вырабатывать перекись водорода и гипохлорит , что уменьшает их противомикробные свойства. Дети, страдающие от таких наследственных нарушений, сильнее подвержены бактериальным инфекциям . Наследственная , связанная с мутациями в гене , кодирующем миелопероксидазу, рассматривается как первичный иммунодефицит .
Недостаточная или избыточная активность ряда белков, входящих в состав гранул нейтрофилов, приводит к болезненным состояниям. При наследственной болезни, известной как , эластаза в составе гранул нейтрофилов недостаточно ингибируется , что приводит к сильнейшим повреждениям тканей при воспалительных процессах, в частности, при хронической обструктивной болезни лёгких . Повышенная активность эластазы нейтрофилов может привести к разрушению лёгочного барьера и острому респираторному дистресс-синдрому . Эластаза нейтрофилов влияет на активность макрофагов, разрушая их Toll-подобные рецепторы и подавляя синтез цитокинов через ингибирование перемещения в ядро транскрипционного фактора NF-κB . При периодической болезни мутации в гене , кодирующем белок пирин, экспрессирующийся преимущественно в нейтрофилах, могут привести к хроническому острому воспалению, что выражается в лихорадке , артралгии , перитоните и, в конце концов, может стать причиной амилоидоза . Имеются данные о связи сниженной активности нейтрофилов, связанной с нарушениями активности миелопероксидазы и дегрануляции, с гипергликемией .
Нейтрофилы связаны с развитием многих хронических воспалительных заболеваний, в том числе аутоиммунных. Они поддерживают хроническое воспаление при системной красной волчанке , псориазе , , АНЦА-ассоциированном системном васкулите , связанном с образованием аутоантител к компонентам цитоплазмы нейтрофилов, ревматоидном артрите , идиопатическом юношеском артрите и (от англ. pyogenic arthritis-pyoderma gangrenosum-acne syndrome ) .
История изучения

Впервые нейтрофилы описал французский исследователь Жан-Батист Сенак , который в 1749 году обнаружил белые клетки в гное. В 1843 году английский врач заметил, что белые клетки в составе гноя происходят от лейкоцитов, которые просочились через кровеносные сосуды. В 1845 году английский врач и физиолог сообщил, что лейкоз обусловлен накоплением белого гноя в кровеносных сосудах, но уже в 1847 году это представление было опровергнуто Рудольфом Вирховым , который показал, что патологические клетки при лейкозе происходят не из гноя, а из самой крови. Первое точное морфологическое описание различных гранулоцитов, в том числе и нейтрофилов, было выполнено Максом Шульце в 1865 году. Кроме морфологических описаний гранулоцитов, Шульце описал и фагоцитоз . Кроме того, в 1876 году фагоцитоз был отмечен Уильямом Ослером
Фагоцитоз был детально изучен и назван Ильёй Ильичом Мечниковым в 1882 году, когда им же была установлена роль фагоцитов в защите от бактерий . В 1879—1880 годах Пауль Эрлих опубликовал несколько работ, в которых детально описал морфологию белых клеток крови. Окрашивание кислыми и щелочными красителями позволило ему выявить соответственно эозинофилы и базофилы , а также тучные клетки . Впоследствии с помощью окрашивания нейтральными красителями он визуализировал сегментоядерные нейтрофилы и их гранулы (за что нейтрофилы и получили своё название). Первоначально Эрлих назвал нейтрофилы «клетками с полиморфными ядрами». Также он описал некоторые аспекты появления, функционирования и дальнейшей судьбы эозинофилов и нейтрофилов . В 1908 году Пауль Эрлих и Илья Мечников разделили Нобелевскую премию по физиологии и медицине .
Примечания
- ↑ , с. 52.
- ↑ Andryukov B. G. , Bogdanova V. D. , Lyapun I. N. (англ.) // Russian journal of hematology and transfusiology. — 2019. — 4 October ( vol. 64 , no. 2 ). — P. 211—221 . — ISSN . — doi : .
- Actor Jeffrey K. (англ.) // Elsevier's Integrated Review Immunology and Microbiology. — 2012. — P. 7—16 . — ISBN 9780323074476 . — doi : .
- Ermert D. , Niemiec M. J. , Röhm M. , Glenthøj A. , Borregaard N. , Urban C. F. (англ.) // Journal Of Leukocyte Biology. — 2013. — August ( vol. 94 , no. 2 ). — P. 223—236 . — doi : . — .
- , с. 73.
- Witko-Sarsat V. , Rieu P. , Descamps-Latscha B. , Lesavre P. , Halbwachs-Mecarelli L. (англ.) // Laboratory Investigation; A Journal Of Technical Methods And Pathology. — 2000. — May ( vol. 80 , no. 5 ). — P. 617—653 . — doi : . — .
- Nathan C. (англ.) // Nature Reviews. Immunology. — 2006. — March ( vol. 6 , no. 3 ). — P. 173—182 . — doi : . — .
- Jacobs L. , Nawrot T. S. , de Geus B. , Meeusen R. , Degraeuwe B. , Bernard A. , Sughis M. , Nemery B. , Panis L. I. (англ.) // Environmental Health : A Global Access Science Source. — 2010. — 25 October ( vol. 9 ). — P. 64—64 . — doi : . — .
- Waugh D. J. , Wilson C. (англ.) // Clinical Cancer Research : An Official Journal Of The American Association For Cancer Research. — 2008. — 1 November ( vol. 14 , no. 21 ). — P. 6735—6741 . — doi : . — .
- De Larco J. E. , Wuertz B. R. , Furcht L. T. (англ.) // Clinical Cancer Research : An Official Journal Of The American Association For Cancer Research. — 2004. — 1 August ( vol. 10 , no. 15 ). — P. 4895—4900 . — doi : . — .
- Yoo S. K. , Starnes T. W. , Deng Q. , Huttenlocher A. (англ.) // Nature. — 2011. — 20 November ( vol. 480 , no. 7375 ). — P. 109—112 . — doi : . — .
- Barer M.R. (англ.) // Medical Microbiology. — 2012. — P. 168—173 . — ISBN 9780702040894 . — doi : .
- Reich D. , Nalls M. A. , Kao W. H. , Akylbekova E. L. , Tandon A. , Patterson N. , Mullikin J. , Hsueh W. C. , Cheng C. Y. , Coresh J. , Boerwinkle E. , Li M. , Waliszewska A. , Neubauer J. , Li R. , Leak T. S. , Ekunwe L. , Files J. C. , Hardy C. L. , Zmuda J. M. , Taylor H. A. , Ziv E. , Harris T. B. , Wilson J. G. (англ.) // PLoS Genetics. — 2009. — January ( vol. 5 , no. 1 ). — P. e1000360—1000360 . — doi : . — .
- ↑ . Дата обращения: 20 сентября 2020. 1 октября 2020 года.
- Niemiec M. J. , De Samber B. , Garrevoet J. , Vergucht E. , Vekemans B. , De Rycke R. , Björn E. , Sandblad L. , Wellenreuther G. , Falkenberg G. , Cloetens P. , Vincze L. , Urban C. F. (англ.) // Metallomics : Integrated Biometal Science. — 2015. — June ( vol. 7 , no. 6 ). — P. 996—1010 . — doi : . — .
- Edwards Steven W. Biochemistry and physiology of the neutrophil. — Cambridge University Press, 1994. — P. 6. — ISBN 978-0-521-41698-6 .
- , с. 53.
- Chu H. T. , Lin H. , Tsao T. T. , Chang C. F. , Hsiao W. W. , Yeh T. J. , Chang C. M. , Liu Y. W. , Wang T. Y. , Yang K. C. , Chen T. J. , Chen J. C. , Chen K. C. , Kao C. Y. (англ.) // BMC Medical Genomics. — 2013. — 12 September ( vol. 6 ). — P. 31—31 . — doi : . — .
- , с. 176.
- ↑ , p. 243.
- ↑ , с. 52—53.
- Zucker-Franklin D, Greaves MF, Grossi CE, Marmont AM. Neutrophils // Atlas of Blood Cells: Function and Pathology. — 2nd. — Philadelphia : Lea & Ferbiger, 1988. — Vol. 1. — P. 168—170. — ISBN 978-0-8121-1094-4 .
- ↑ , с. 177—178.
- , с. 54—55.
- , с. 54.
- ↑ , с. 55.
- ↑ , с. 75.
- , с. 89.
- , с. 201—204.
- Tak T. , Tesselaar K. , Pillay J. , Borghans J. A. , Koenderman L. (англ.) // Journal Of Leukocyte Biology. — 2013. — October ( vol. 94 , no. 4 ). — P. 595—601 . — doi : . — .
- Pillay J. , den Braber I. , Vrisekoop N. , Kwast L. M. , de Boer R. J. , Borghans J. A. , Tesselaar K. , Koenderman L. (англ.) // Blood. — 2010. — 29 July ( vol. 116 , no. 4 ). — P. 625—627 . — doi : . — .
- Wheater Paul R., Stevens Alan. . — Edinburgh : Churchill Livingstone, 2002. — ISBN 978-0-443-07001-3 .
- ↑ , с. 91.
- ↑ , p. 244.
- ↑ , с. 56.
- Tan S. Y. , Weninger W. (англ.) // Current Opinion In Immunology. — 2017. — February ( vol. 44 ). — P. 34—42 . — doi : . — .
- Lämmermann T. (англ.) // Journal Of Leukocyte Biology. — 2016. — July ( vol. 100 , no. 1 ). — P. 55—63 . — doi : . — .
- Chtanova T. , Schaeffer M. , Han S. J. , van Dooren G. G. , Nollmann M. , Herzmark P. , Chan S. W. , Satija H. , Camfield K. , Aaron H. , Striepen B. , Robey E. A. (англ.) // Immunity. — 2008. — 19 September ( vol. 29 , no. 3 ). — P. 487—496 . — doi : . — .
- Henry K. M. , Loynes C. A. , Whyte M. K. , Renshaw S. A. (англ.) // Journal Of Leukocyte Biology. — 2013. — October ( vol. 94 , no. 4 ). — P. 633—642 . — doi : . — .
- , p. 243—244.
- Brinkmann V. , Reichard U. , Goosmann C. , Fauler B. , Uhlemann Y. , Weiss D. S. , Weinrauch Y. , Zychlinsky A. (англ.) // Science (New York, N.Y.). — 2004. — Vol. 303, no. 5663 . — P. 1532—1535. — doi : . — .
- Urban C. F. , Ermert D. , Schmid M. , Abu-Abed U. , Goosmann C. , Nacken W. , Brinkmann V. , Jungblut P. R. , Zychlinsky A. (англ.) // PLoS Pathogens. — 2009. — October ( vol. 5 , no. 10 ). — P. e1000639—1000639 . — doi : . — .
- Clark S. R. , Ma A. C. , Tavener S. A. , McDonald B. , Goodarzi Z. , Kelly M. M. , Patel K. D. , Chakrabarti S. , McAvoy E. , Sinclair G. D. , Keys E. M. , Allen-Vercoe E. , Devinney R. , Doig C. J. , Green F. H. , Kubes P. (англ.) // Nature Medicine. — 2007. — April ( vol. 13 , no. 4 ). — P. 463—469 . — doi : . — .
- Gupta A. K. , Hasler P. , Holzgreve W. , Hahn S. (англ.) // Seminars In Immunopathology. — 2007. — June ( vol. 29 , no. 2 ). — P. 163—167 . — doi : . — .
- Hoyer F. F. , Nahrendorf M. (англ.) // European Heart Journal. — 2017. — 14 February ( vol. 38 , no. 7 ). — P. 465—472 . — doi : . — .
- Mangold A. , Alias S. , Scherz T. , Hofbauer T. , Jakowitsch J. , Panzenböck A. , Simon D. , Laimer D. , Bangert C. , Kammerlander A. , Mascherbauer J. , Winter M. P. , Distelmaier K. , Adlbrecht C. , Preissner K. T. , Lang I. M. (англ.) // Circulation Research. — 2015. — 27 March ( vol. 116 , no. 7 ). — P. 1182—1192 . — doi : . — .
- Azzouz Dhia , Palaniyar Nades. (англ.) // Cell Death & Disease. — 2018. — August ( vol. 9 , no. 8 ). — ISSN . — doi : .
- Gerasimov I. G. , Ignatov D. Iu. // Tsitologiia. — 2001. — Т. 43 , № 5 . — С. 432—436 . — .
- Gerasimov I. G. , Ignatov D. Iu. // Tsitologiia. — 2004. — Т. 46 , № 2 . — С. 155—158 . — .
- Gerasimov I. G. , Ignatov D. Iu. , Kotel'nitskiĭ M. A. // Tsitologiia. — 2005. — Т. 47 , № 6 . — С. 549—553 . — .
- Gerasimov I. G. , Ignatov D. Iu. // Tsitologiia. — 2005. — Т. 47 , № 6 . — С. 554—558 . — .
- , с. 90.
- Tecchio C. , Micheletti A. , Cassatella M. A. (англ.) // Frontiers In Immunology. — 2014. — Vol. 5 . — P. 508—508 . — doi : . — .
- ↑ Silvestre-Roig C. , Fridlender Z. G. , Glogauer M. , Scapini P. (англ.) // Trends In Immunology. — 2019. — July ( vol. 40 , no. 7 ). — P. 565—583 . — doi : . — .
- . Дата обращения: 20 сентября 2020. 25 ноября 2020 года.
- Al-Gwaiz L. A. , Babay H. H. (англ.) // Medical Principles And Practice : International Journal Of The Kuwait University, Health Science Centre. — 2007. — Vol. 16 , no. 5 . — P. 344—347 . — doi : . — .
- ↑ Mitchell, Richard Sheppard, Kumar, Vinay, Abbas, Abul K., Fausto, Nelson. Robbins Basic Pathology. — Philadelphia : Saunders. — ISBN 1-4160-2973-7 .
- . Дата обращения: 10 апреля 2013. Архивировано из 21 мая 2013 года.
- . omim.org . Дата обращения: 17 марта 2020. 13 марта 2020 года.
- Kawabata K. , Hagio T. , Matsuoka S. (англ.) // European Journal Of Pharmacology. — 2002. — 6 September ( vol. 451 , no. 1 ). — P. 1—10 . — doi : . — .
- Domon H. , Nagai K. , Maekawa T. , Oda M. , Yonezawa D. , Takeda W. , Hiyoshi T. , Tamura H. , Yamaguchi M. , Kawabata S. , Terao Y. (англ.) // Frontiers In Immunology. — 2018. — Vol. 9 . — P. 732—732 . — doi : . — .
- Ozen S. (англ.) // European Journal Of Pediatrics. — 2003. — July ( vol. 162 , no. 7-8 ). — P. 449—454 . — doi : . — .
- Xiu F. , Stanojcic M. , Diao L. , Jeschke M. G. (англ.) // International Journal Of Endocrinology. — 2014. — Vol. 2014 . — P. 486403—486403 . — doi : . — .
- ↑ Kay A. B. (англ.) // Microbiology Spectrum. — 2016. — August ( vol. 4 , no. 4 ). — doi : . — .
- Ambrose C. T. (англ.) // Cellular Immunology. — 2006. — March ( vol. 240 , no. 1 ). — P. 1—4 . — doi : . — .
- Gordon S. (англ.) // Immunity. — 2016. — 15 March ( vol. 44 , no. 3 ). — P. 463—475 . — doi : . — .
- . Дата обращения: 20 сентября 2020. 14 августа 2018 года.
Литература
- Афанасьев Ю. И., Кузнецов С. Л., Юрина Н. А., Котовский Е. Ф. и др. Гистология, цитология и эмбриология. — 6-е изд., перераб. и доп.. — М. : Медицина, 2004. — 768 с. — ISBN 5-225-04858-7 .
- Руководство по гематологии: в 3 т. / Под ред. А. И. Воробьёва. — М. : Ньюдиамед, 2002. — Т. 1. — 280 с. — ISBN 5-88107-038-0 .
- Хаитов Р. М. Иммунология: структура и функции иммунной системы. — М. : ГЭОТАР-Медиа, 2019. — 328 с. — ISBN 978-5-9704-4962-2 .
- Ярилин А. А. Иммунология. — М. : ГЭОТАР-Медиа, 2010. — 752 с. — ISBN 978-5-9704-1319-7 .
- Anthony L. Mescher. . — McGraw-Hill Education, 2016. — ISBN 978-0-07-184270-9 .
| Кроветворение | |
|---|---|
| Компоненты | |
| Биохимия | |
| Заболевания | |
|
См. также:
Гематология
,
Онкогематология
|
|
| Органы | |||||
|---|---|---|---|---|---|
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
- 2020-01-19
- 1