Обыкновенные губки
- 1 year ago
- 0
- 0

Гу́бки ( лат. Porifera ) — тип водных, преимущественно морских, многоклеточных животных , ведущих прикреплённый образ жизни. Распространены по всему земному шару от прибрежной зоны и до почти максимальных глубин океана. Насчитывают около 8000 видов.
Губки не имеют настоящих тканей и органов , и различные функции выполняют разнообразные отдельные клетки и клеточные пласты. Питание большинства видов осуществляется путём фильтрации воды, прогоняемой через расположенную внутри тела губки водоносную систему различной сложности. Изнутри значительную часть водоносной системы выстилают особые клетки со жгутиком и воротничком из микроворсинок — хоаноциты , составляющие в совокупности хоанодерму; остальная часть водоносной системы и внешние покровы образованы пластом пинакоцитов (пинакодермой). Между пинакодермой и хоанодермой расположен — слой внеклеточного матрикса , содержащий разнообразные по строению и выполняемым функциям отдельные клетки. У многих губок в мезохиле обитают эндосимбиотические прокариоты ( бактерии и археи ), а также эукариотические водоросли.
Размножаются половым и бесполым путями (в том числе с образованием покоящихся почек — геммул ). Обладают высокой способностью к регенерации . Многие губки, а также населяющие их микроорганизмы вырабатывают биологически активные вещества , используемые в медицине .

Аристотель (IV век до н. э.), стоявший у истоков научной систематики животных, разделил их в своём сочинении « О частях животных » ( др.-греч. Περὶ ζώων μορίων ) на две большие группы — «животные, обладающие кровью» и «животные без крови», выделив в каждой из них более мелкие подразделения. Среди подразделений второй из этих групп (примерно соответствующей позднейшим беспозвоночным ) он, в частности, выделил «безраковинных свободноживущих» ( голотурии , гидроиды ) и «безраковинных прирастающих» ( актинии , губки), трактуя их как организмы, в которых смешана животная и растительная природа .
Позднее (в IV—V веках н. э.) неоплатоники ( Дексипп , Аммоний Гермий ) стали называть такие организмы « зоофитами » ( др.-греч. ζωόφυτα ), причисляя их к промежуточным формам между растениями и животными. В средневековой Европе данный термин почти не применялся, но вновь вошёл в употребление в эпоху Возрождения : его применяли в своих классификациях такие зоологи, как Эдвард Уоттон , Гийом Ронделе , Конрад Геснер и Улиссе Альдрованди . Губки неизменно фигурировали в составе «зоофитов», хотя объём группы менялся; так, в сочинении Уоттона « De differentiis animalium » (1552), первом обстоятельном зоологическом трактате со времён Аристотеля, к зоофитам были добавлены также морские звёзды , сцифоидные медузы и гребневики . В книге Карла Линнея « Systema Naturae » в состав класса Vermes вошёл отряд Zoophyta с ещё более широким, чем у Уоттона, объёмом: Линней включил в число зоофитов также морских ежей и некоторых моллюсков и червей . При этом в 10-м издании книги (1758) Линней определял Zoophyta как «растения с цветами, живущими животной жизнью», а в 12-м издании (1766—1768) — как «сложных животных, расцветающих подобно цветам растений» .

Животная природа губок впервые была обоснована Джоном Эллисом , который описал их способность создавать ток воды и изменять диаметр и изложил свои наблюдения в 1765 году в письме к Даниэлю Соландеру (в 1766 году письмо было опубликовано в Философских трудах Королевского общества ) . В 1752 году, до открытия Эллиса, французский естествоиспытатель высказывал гипотезу, согласно которой губки являются не живыми организмами, а конструкциями, возводимыми стрекающими или морскими червями .
Систематическое положение губок среди «зоофитов» постепенно уточнялось. У Жана-Батиста Ламарка в сочинениях « Système des animaux sans vertèbres » (1801) и « Philosophie zoologique » (1809) губки фигурируют в составе отряда «Полипы с полипняком» — вместе со мшанками , оболочниками и рядом групп кишечнополостных . Жорж Кювье в своём сочинении « Le régne animal distribué d’apres son organisation » (1817) включил губок вместе с коралловыми полипами в класс Polypae раздела Radiata (последний примерно соответствует Zoophyta в смысле Уоттона или Линнея, хотя Кювье уже не считал своих Radiata переходными между растениями и животными организмами) .

Переход к углублённому изучению биологии губок наметился в исследованиях , в которых он предложил первую точную интерпретацию строения и физиологии данных животных. Уже в первой своей статье (1825) Грант описал личинок и яйца у губок из рода , провёл тщательное микроскопическое исследование структуры пор губок . В 1836 году он ввёл для губок название Porophora , которое было заменено самим же Грантом в 1841 году на Poriphera , а в 1847 году — на Porifera (все названия означают «несущие поры») .
На протяжении большей части XIX века губок обычно сближали с кишечнополостными и часто просто включали в состав последних (хотя такие естествоиспытатели, как Рене Дютроше , Поль Жерве и Джон Хогг , относили губок к растениям ). Однако ещё в 1816 году Анри де Блэнвиль выдвинул идею о тесной связи губок с простейшими (Protozoa) , а в 1860 году Хогг и в 1866 году Эрнст Геккель предложили выделить отдельное царство (у Хогга — Protoctista, у Геккеля — Protista ), в которое включили губок, одноклеточных животных и часть одноклеточных растений ( Томас Гексли же, считавший губок колониями одноклеточных, в 1864 году попросту включил их в состав Protozoa) . Подобная точка зрения нашла, казалось, своё подтверждение в открытии в 1867 году воротничковых жгутиконосцев ( хоанофлагеллят ), удивительно сходных с имеющимися у всех губок особыми клетками — хоаноцитами .

Однако последняя треть XIX века стала, по выражению А. В. Ересковского, «золотым веком» эмбриологии губок (и первым этапом систематических исследований их развития) . В 1874—1879 годах исследования И. И. Мечникова , Франца Шульце и Оскара Шмидта , посвящённые изучению строения и развития губок, неопровержимо доказали их принадлежность к многоклеточным животным. Одновременно было обнаружено большое своеобразие данной группы животных. В частности, Шульце (1878) и Ив Делаж (1892) описали так называемое явление «извращения зародышевых листков» при развитии губок, резко отличающее Porifera от всех других Metazoa (включая кишечнополостных); впрочем, в конце XX — начале XXI века терминология изменилась: стало преобладать мнение, по которому у губок зародышевые листки в ходе эмбриогенеза вообще не формируются, а потому об их «извращении» говорить нет смысла . Поэтому Уильям Соллас (1884) противопоставил губок как Parazoa всем остальным Metazoa (получившим вскоре название Eumetazoa ). В поздней системе Геккеля (1894—1895) губки уже выведены из состава Protista и рассматриваются в царстве животных как самостоятельный тип Spongiae, а в системе Рэя Ланкестера (1900—1909) они со всей определённостью отнесены к Metazoa и фигурируют как тип Porifera (единственный в разделе Parazoa). Эта последняя точка зрения и являлась абсолютно преобладающей на протяжении почти всего XX века, хотя ранг Parazoa у разных авторов варьирует: то это «раздел», то — «надраздел», то — « подцарство » .
В 1900—1960 годах (второй этап исследований развития губок, по А. В. Ересковскому) интерес к изучению развития губок снижается, хотя и появляются важные работы и . Примерно в 1960 году начинается третий этап, для которого характерно преобладание ультраструктурных исследований с применением электронной микроскопии . В конце XX века началось изучение цитогенетических и молекулярно-биологических особенностей губок .
— раздел зоологии изучающий губки.

Основание тела губок прочно прикрепляется к , благодаря чему они неподвижны. В связи с этим в их теле прослеживается апикобазальная полярность: от нижней (базальной) части тела, прикреплённой к субстрату, до верхней (апикальной). Характерная для остальных Metazoa передне-задняя полярность у губок не выражена . Размеры тела представителей группы по апикобазальной оси варьируют в широких пределах: от нескольких миллиметров до 1,5—2 м . Самой крупной губкой в настоящее время считается неидентифицированный вид шестилучевых губок из семейства (найден на глубине 2,1 км вблизи Гавайских островов ); она достигает 3,5 м в длину, 2 м в высоту и 1,5 м в ширину . Среди колониальных видов наибольшие размеры известны для колонии губок , найденной у западной части побережья Канады и достигающей 3,4 м в длину, 1,1 м в высоту и 0,5 м в ширину . Губки отличаются неограниченным ростом: верхний предел увеличения размеров тела, характерный для большинства видов животных, у них фактически отсутствует .
Форма тела губок чрезвычайно разнообразна. Они могут иметь вид корковых, подушковидных, плёнковидных, комкообразных обрастаний и наростов на субстрате . Среди них известны шаровидные, цилиндрические, бокаловидные, воронковидные, трубчатые, стебельчатые, ветвистые, вееровидные и иные формы . В 1964 году в Антарктике на глубине около 4 км был обнаружен объект странной формы, позднее известный как антенна Элтанина . Сторонники НЛО признали его артефактом внеземного происхождения, однако, по мнению большинства учёных, на самом деле это — редкая хищная губка Chondrocladia concrescens . Поверхность тела губок обычно является неровной: игольчатой или даже щетинистой, и лишь в редких случаях она может быть относительно гладкой и ровной. Одни виды губок имеют мягкое и эластичное тело, другие — более жёсткое или твёрдое .
Окраска тела губок также разнообразна. Многие виды благодаря наличию клеточных пигментов либо эндосимбионтов ярко окрашены: преобладают жёлтый, красный, коричневый, оранжевый, зелёный, фиолетовый цвета и их различные оттенки . Лишённые пигментов губки характеризуются белой или серой окраской .

В теле губок невозможно выделить настоящие ткани, органы и их системы , присущие другим животным. Таким образом, губки взаимодействуют с окружающей средой не при помощи обособленных многоклеточных систем, а единым образом во всех частях своего тела. Через небольшие отверстия на поверхности тела ( , или поры) вода проникает в тело губки и далее циркулирует по связанной системе полостей и каналов (приводящих и отводящих), которые заканчиваются одним или несколькими крупными отверстиями — . Самый крупный канал, переходящий в оскулюм, называется атриумом, или . Водоносная система позволяет губке каждые 5 секунд пропускать через себя объём воды, равный собственному, чтобы снабжать клетки питательными веществами и кислородом . Кроме того, с током воды губка выводит из своего тела токсичные вещества и продукты метаболизма, а также гаметы . Губки постоянно перестраивают свою водоносную систему, повышая её эффективность, чтобы с большой эффективностью фильтровать воду в изменяющихся гидродинамических условиях .

Поверхность тела губки, а также каналы водоносной системы образованы уплощёнными, плотно прилегающими друг к другу клетками — пинакоцитами . Расширенные участки каналов, именуемые хоаноцитными, или жгутиковыми камерами, выстланы хоаноцитами — клетками, снабжёнными жгутиком и способными к фильтрации пищевых частиц и созданию тока воды в водоносной системе. Входное отверстие хоаноцитной камеры называется прозопиль, а выходное — апопиль. Слои пинакоцитов и хоаноцитов разделены — внутренним пространством губки, которое непосредственно не сообщается с водой . Подробно строение и состав клеток губок будут рассмотрены ниже.
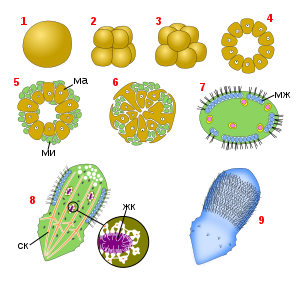
Простейший вариант строения губок — асконоидный, при котором имеются обширная хоаноцитная выстилка мезохила и наружный слой пинакоцитов, который связан со слоем хоаноцитов узкими проводящими каналами; хоаноцитная полость открывается непосредственно оскулярным отверстием. Асконоидных губок иногда называют гомоцельными, потому что водоносная система у них выстлана только хоаноцитами. У видов с более сложным строением некоторые участки водоносной системы сформированы пинакоцитами, а не хоаноцитами, и такие губки называются гетероцельными . Асконоидные губки обычно имеют вид соединяющихся ( анастомозирующих ) трубочек, не превышая в диаметре 1 мм. Например, у представителей рода такие трубочки соединяются основаниями и собраны в пучки, а у тело представлено трубчатой сетью . Для увеличения размера тела при сохранении асконоидной организации можно было бы увеличивать размер атриальной полости и/или объём мезохила. Однако ни то, ни другое невозможно по физическим причинам: при слишком большом объёме атриальной полости в ней происходил бы застой воды, так как оптимальное соотношение площади поверхности и объёма было бы нарушено и площади поверхности хоанодермы не хватало бы для эффективной прогонки воды, а чрезмерное увеличение объёма мезохила помешало бы поверхностному слою клеток адекватно обслуживать протекающие там процессы .

Тем не менее, размер большинства губок значительно превосходит размеры асконоидных форм. Для решения описанных выше физических проблем слои пинакоцитов и хоаноцитов изгибаются навстречу друг другу, причём хоаноциты группируются в карманообразных камерах, которые одним концом открываются в атриум (выстланный уже не хоаноцитами, а пинакоцитами), а другим сообщаются с внешней средой посредством коротких приводящих каналов. Такой тип организации водоносной системы губок называется сиконоидным . Сиконоидное строение водоносной системы позволяет увеличить площадь поверхности и одновременно уменьшить объём атриума. В такой водоносной системе вода проделывает следующий путь: остии → приводящие каналы → прозопили → жгутиковые каналы → атриум → оскулюм. Сиконоидные губки обычно крупнее асконоидных, их диаметр может составлять от одного до нескольких сантиметров. Примерами губок с сиконоидным строением водоносной системы могут служить представители родов и .
Дальнейшее усложнение строения губки происходит при сглаживании поверхности наружного пинакоцитного слоя и появлении под пинакодермой широких полостей, сообщающихся с внешней средой через узкие поры. Радиально расположенные хоаноцитные камеры открываются уже не в атриум, а в выстланную пинакоцитами полость, которая и сообщается с атриумом . В этом случае говорят о силлеибидном типе водоносной системы. Такая водоносная система имеется у многих Homoscleromorpha .
Увеличение количества хоаноцитных камер, уменьшение их размеров, погружение их в пространство между стенкой атриальной полости и наружным слоем пинакоцитов, развитие сложной сети приводящих и отводящих каналов приводят к созданию лейконоидной водоносной системы, которая позволяет губкам достигать максимально возможных размеров 1—2 м . Жгутиковые камеры у лейконоидных губок очень многочисленны; например, у Microciona porifera на 1 мм³ приходится около 10000 камер, каждая из которых имеет диаметр от 20 до 39 мкм и содержит около 57 хоаноцитов. Лейконоидная организация присуща большинству морских губок, населяющих мелководье, и всем пресноводным губкам. Лейконоидные губки могут иметь самую разнообразную форму, даже быть похожими на вазы .
В 2011 году для описания водоносной системы губок рода Leucascus был введён новый термин — соленоидная водоносная система. Она характеризуется тем, что хоаноциты располагаются только в анастомозирующих трубочках, а пинакоциты выстилают атриальную полость. Таким образом, соленоидная водоносная система также является гетероцельной .
В ходе развития губок их водоносная система может останавливаться на начальной асконоидной стадии либо развиваться до одного из описанных выше более сложных типов — сиконоидного, силлеибидного или лейконоидного .
В случае асконоидного типа строения пространство атриальной полости, выстланное хоаноцитами, можно считать одной большой хоаноцитной камерой, у которой имеется много прозопилей, соответствующих внутренним отверстиям приводящих каналов, и один апопиль, совпадающий с оскулюмом. При трёх других более сложных типах строения в теле губки можно выделить зону, в которой располагаются хоаноцитные камеры — хоаносому. У некоторых губок между хоаносомой и наружной пинакодермой находится слой мезохила, в котором хоаноцитных камер нет; его называют эктосома. Иногда к пинакоцитам примыкает более плотный тонкий слой мезохила, и эту самую внешнюю часть губок называют кортикальным слоем, или кортексом. В базальной части губок хоаноцитных камер, как правило, нет .
Впрочем, все типы водоносной системы встречаются только у известковых губок, в то время как у обыкновенных и стеклянных губок, а также у Homoscleromorpha имеется множество вариантов лейконоидной организации .
Проблема индивидуальности или колониальности губок не до конца решена. Во второй половине XX века было предложено рассматривать губок в качестве модульных организмов, состоящих из водоносных модулей. Под модулем понимают определённый объём губки, обслуживаемый системой хоаноцитных камер и водоносных каналов, связанных с одним оскулюмом .

Губки не имеют настоящих тканей, в отличие от большинства животных. Вся их жизнедеятельность обеспечивается активностью отдельных клеток и клеточных пластов, имеющих разное строение и функции. Однако в отличие от настоящих тканей клетки клеточных слоёв, хотя и выполняют сходные функции, имеют разное происхождение . Внешний и внутренний клеточные слои — пинакодерма и хоанодерма, а также мезохил, представляющий собой совокупность клеток и внеклеточных элементов, в структурном и функциональном плане устроены существенно проще, чем их аналоги у других Metazoa. Многие клетки губок могут претерпевать в клетки других типов. У губок нет единой категории тотипотентных или плюрипотентных клеток. У обыкновенных губок их функции выполняют и хоаноциты, у известковых губок — хоаноциты, у Homoscleromorpha — хоаноциты и пинакоциты, у стеклянных губок , вероятно, археоциты. Стеклянные губки отличаются от остальных губок синцитиальной организацией, то есть большая часть их клеток слиты в единую сеть; строение синцития стеклянных губок будет описано отдельно. Ниже будет рассмотрена организация губок, имеющих клеточное строение .
Пинакодерма подразделяется на экзопинакодерму, покрывающую тело губки снаружи, базипинакодерму, которая развивается в основании губки и прикрепляет её к субстрату, и эндопинакодерму, образующую стенки внутренних полостей и каналов водоносной системы .
Экзопинакодерма тонка и эластична и покрыта протеогликановым матриксом . Степень развития экзопинакодермы различается у разных губок. Иногда она подстилается кортексом, который, помимо клеток и спикул, может иногда содержать даже инородные включения. У некоторых губок один слой экзопинакоцитов подстилает мощный мезохил, в то время как у других экзопинакодерму толщиной 3 мкм подстилает почти такой же тонкий мезохил . Экзопинакоциты — покровные клетки губок, которые снаружи выглядят как уплощённые многоугольники , похожие на чешуйки и плотно примыкающие друг к другу, образуя своего рода клеточную «брусчатку» (разделяющий мембраны соседних экзопинакоцитов межклеточный зазор обычно составляет лишь 15 нм ). Экзопинакоциты лишены жгутиков , но у некоторых Homoscleromorpha они несут реснички . Возможно, у большинства видов между экзопинакоцитами нет специализированных межклеточных контактов , однако они связаны хорошо развитой адгезионной системой гликокаликса . Экзопинакоциты могут синтезировать и выводить на свою поверхность компоненты межклеточного матрикса, в том числе коллаген .
В экзопинакодерме находятся поры — многочисленные отверстия диаметром от 4 до 100 мкм , через которые вода проникает в водоносную систему губки. У большей части обыкновенных губок и всех Homoscleromorpha поры — это межклеточные отверстия, а у известковых губок поры формируются внутри особых цилиндрических трубчатых клеток — . В последнем случае диаметр поры регулируется сокращением цитоплазматических волокон, поэтому пороциты можно рассматривать как маленькие клапаны, работающие по принципу сфинктеров . У некоторых губок пороциты способны к сокращению в ответ на механическое или химическое раздражение. Описаны обыкновенные губки, у которых есть уплощённые пороциты с отверстием в центре и на периферии, способным открываться или закрываться .
Экзопинакодерма выполняет многие функции, присущие эпителиям эуметазоев — такие, как абсорбция , секреция, транспорт, выделение, защита . С эпителиями других животных пинакодерму сближает наличие десмосом, которые есть по крайней мере у части губок. У некоторых пресноводных губок между клетками пинакодермы, вероятно, есть и плотные контакты . Возможно, между пинакоцитами существуют и щелевые контакты (предполагается, что пинакодерма некоторых губок способна к медленному проведению электрических сигналов). Медленное (около 0,25 см/c) проведение электрических импульсов открыто в 1990-е годы у стеклянных губок. Импульсы распространяются по трабекулярному синцитию — особой ткани .
Базипинакоциты — уплощённые клетки, формирующие базальную поверхность губок и обеспечивающие прикрепление к субстрату за счёт синтеза и хитина . У обыкновенных губок, имеющих массивный арагонитовый скелет, базипинакоциты участвуют в синтезе карбоната кальция . Некоторые губки способны к медленному (4-5 мм в сутки) ползанию. При локомоции образуется лидирующий край из подвижных клеток базипинакодермы .
Эндопинакоциты выстилают приносящие и выносящие каналы водоносной системы и представляют собой уплощённые, веретеновидные на срезах клетки. Их подразделяют на прозопинакоциты, образующие приносящие каналы, и апопинакоциты, образующие выносящие каналы. Эндопинакоциты закрепляются во внеклеточном матриксе при помощи псевдоподиальных выростов, образуемых базальной частью клеток. Между собой они контактируют путём простого прилегания .
У некоторых обыкновенных губок в пинакодерме имеются сократимые клетки — миоциты . Это длинные веретеновидные клетки со скоплениями сократимых волокон в цитоплазме. У пресноводных губок пусти актиновых филаментов залегают не в миоцитах, а в эндопинакоцитах оскулярных трубок .
В отличие от эпителиальных клеток эуметазоев пинакоциты губок могут сокращаться, принимать амёбовидную форму, перемещаться и фагоцитировать пищевые частицы, выводя непереваренные остатки наружу .

Хоанодерма образована клетками одного типа — хоаноцитами, снабжёнными жгутиком, окружённым воротничком микроворсинок (микровиллей). Длина микровиллей составляет 2/3 длины жгутика. В отличие от пинакоцитов, хоаноциты могут быть цилиндрическими, кубическими, трапециевидными или немного уплощёнными. Хоаноциты — очень маленькие клетки, достигают всего 2—10 мкм в диаметре. Микровилли соединены между собой нитями протеогликанов ( гликокаликс ). Жгутик хоаноцита отходит от небольшого бугорка или кольцевидного выпячивания на поверхности клетки. Его аксонема имеет типичное для эукариот строение. В зависимости от условий обитания губки, её физиологического состояния или стадии онтогенеза форма хоаноцитов может варьировать . Биение жгутика хоаноцита создаёт область низкого давления у его основания, и туда устремляется вода, попутно проходя через воротничок микровиллей и оставляя на них пищевые частицы. Далее вода движется вдоль жгутика хоаноцита и покидает его .
Хоаноциты объединяются в хоаноцитные (жгутиковые) камеры, которые у обыкновенных губок подстилаются внеклеточным матриксом, содержащим коллаген. У Homoscleromorpha хоанодерма, как и пинакодерма, подстилается настоящей базальной пластинкой с коллагеном IV типа. У обыкновенных губок в одной жгутиковой камере находится 50—200 хоаноцитов, у стеклянных губок — 200—300, а у сиконоидных известковых губок — до 1000. В хоаноцитной камере микровилли и жгутики каждого хоаноцита направлены к выходу из камеры. Проходящая через камеру вода заходит через микровилли к основанию каждого жгутика, далее выходит в полость камеры и покидает её .
Хоаноциты — одни из наиболее хорошо изученных клеток губок. Они могут дифференцироваться в сперматоциты и даже ооциты . Некоторые исследования показали, что хоаноциты могут превращаться и в археоциты, поэтому их можно рассматривать как стволовые клетки . Кругооборот хоаноцитов происходит очень быстро. Они заменяются быстрее, чем какие-либо другие клетки в теле губки .
На границе хоаноцитной камеры и выносящего канала располагаются особые апопилярные клетки, переходные между апопинакоцитами и хоаноцитами. Считается, что они формируются из хоанобластов на ранних этапах формирования жгутиковой камеры . Они имеют треугольную форму на поперечном срезе, а их края, обращённые в камеру, снабжены гребнем микровиллей. Апопилярные клетки описаны у некоторых обыкновенных губок и всех Homoscleromorpha. Предполагается, что они регулируют поток в водоносной системе .
Прозопили (входные отверстия хоаноцитных камер) могут быть образованы псевдоподиальными выростами хоаноцитов или пинакоцитами, которые контактируют с хоаноцитами и образуют что-то наподобие пороцита. Апопили (выходные отверстия) крупнее прозопилей и могут быть образованы либо апопилярными клетками, либо особыми конусовидными клетками. В отличие от хоаноцитов, имеющих один жгутик, конусовидные клетки снабжены ресничками , которые медленно бьются по направлению из камеры. У некоторых губок, например, Tethya wilhelma , конусовидных клеток нет, и вместо этого несколько клеток, похожих на эндопинакоциты, образуют ситовидное поле на выходе из камеры .
У многих обыкновенных губок в хоаноцитных камерах имеются особые центральные клетки. У некоторых видов центральная клетка представляет собой структуру неправильной формы, располагающуюся на выходе из жгутиковой камеры и снабжённую обширным каналом, в который входят жгутики хоаноцитов. В других случаях центральная клетка имеет вид перфорированной пластинки, выстилающей хоаноцитную камеру поверх апикальной части хоаноцитов, причём жгутики проходят в просвет камеры через отверстия в центральной клетке. Предполагаются, что центральные клетки участвуют в регуляции биения жгутиков в пределах одной камеры. Однако центральные клетки есть не во всех хоаноцитных камерах одной губки .

Мезохил — это внутреннее пространство тела губок, отграниченное от внешней среды слоями хоаноцитов и пинакоцитов. Мезохил состоит из скелетных элементов, молекул внеклеточного матрикса и клеток, выполняющих разнообразные функции. В зависимости от выполняемых функций клетки мезохила можно подразделить на две группы: клетки одной группы выполняют опорно-соединительную функцию, а клетки другой группы — защитно-секреторную. Клетки мезохила не образуют скоплений и могут менять форму и передвигаться, из-за чего иногда всю совокупность клеток мезохила называют амёбоцитами . Для каждой таксономической группы губок характерна определённая совокупность типов клеток мезохила, обладающая уникальными функциями и цитологическими особенностями .

Клетки этой группы формируют элементы органического и минерального скелета, а также внеклеточный матрикс. Колленциты — это подвижные клетки, секретирующие коллаген. По сути, колленциты — это разновидность фибробластов . Они имеют звёздчатую или веретеновидную форму и образуют ветвящиеся псевдоподии. Ядро лишено ядрышка , а в цитоплазме нет специфических включений. В ходе экспериментов по пересадке фрагментов тела одной губки к другой губке было показано, что колленциты участвуют в защитных реакциях. В качестве разновидности колленцитов рассматриваются лофоциты — амёбоидные клетки с чётко выраженной передне-задней полярностью, секретирующие элементы внеклеточного матрикса и отвечающие за его организацию в фибриллярные пучки. Внешне лофоциты напоминают археоциты . Передний полюс снабжён короткими звёздчатыми псевдоподиями, а задний конец оставляет за собой след в виде пучка коллагеновых фибрилл, объединённых гликопротеиновым матриксом. Ядро лофоцитов имеет ядрышко, в цитоплазме сильно развит шероховатый эндоплазматический ретикулум , хорошо заметны микротрубочки и микрофиламенты . Главная функция лофоцитов — пополнение и возобновление коллагеновых волокон мезохила . У обыкновенных губок колленциты и лофоциты считаются производными археоцитной линии .

Склероциты секретируют минеральные элементы скелета — спикулы , выполняющие опорную функцию и поддерживающие форму тела губки. У кремниевых губок склероциты осуществляют внутриклеточный синтез спикул. Из всех Metazoa только обыкновенные и стеклянные губки используют для построения скелета кремний , а не кальций . По размеру образуемых спикул выделяют мегасклероцит и микросклероциты. Спикула синтезируется внутри вакуоли , образованной особой мембраной — силикалеммой, которая не контактирует ни с какими мембранами клетки . Склероциты известковых губок происходят из пинакоцитов и, возможно, хоаноцитов .
Кроме внутреннего основного скелета многие губки имеют ещё и кожный (эктосомальный) скелет, защищающий губку от хищников и других губок. Его строение разнообразно и может включать как горизонтально ориентированные спикулы, собранные в стопки, так и вертикально ориентированные спикулы, образующие частокол .
Спонгоциты образуют околоспикулярный спонгин или «роговой» скелет у обыкновенных губок, лишённых минерального скелета. Эти клетки мигрируют к склероцитам, синтезирующим спикулы, или готовым спикулам и прикрепляют к ним молекулы спонгина — особой формы коллагена, содержащей высокую концентрацию йода . Спонгин встречается только у губок и имеет вид толстых скелетных волокон . Считаются, что спонгоциты характеризуются положительным хемотаксисом к кремнию. При образовании органического скелета спонгоциты группируются, соединяясь пальцевидными контактами. Считается, что у обыкновенных губок спонгоциты, как и склероциты, — производные археоцитной линии .
Все клетки этой группы можно условно подразделить на амёбоидные клетки и клетки с включениями. Основные функции этой группы клеток — защитная (фагоцитарная, бактерицидная ), запасающая, секреция основного вещества мезохила, а также передача частичек пищи и кислорода .
У обыкновенных губок имеются типичные амёбоидные клетки — археоциты, отличающиеся большим ядром и ядрышком, а также многочисленными крупными лизосомами , способными к фагоцитозу . У Homoscleromorpha такие клетки есть не у всех видов. В настоящее время под археоцитами понимают как тотипотентные ( стволовые ) клетки, как и любые клетки, имеющие ядро с ядрышком и многочисленные фагосомы . Археоциты — это наиболее активно делящаяся фракция свободных клеток. Они активно участвуют в бесполом и половом размножении, а также восстановительном морфогенезе .
Бактериоциты — это подвижные амёбоидные клетки с вакуолями, в которых обитают прокариотические эндосимбионты. Пока они выявлены только у некоторых видов обыкновенных губок. Некоторые бактериоциты содержат внутри одну большую вакуоль с бактериями, а другие имеют многочисленные мелкие вакуоли, в которых заключены одна или несколько бактерий. Бактериоциты могут быть довольно крупными (до 20 мкм). У некоторых видов бактериоциты осуществляют вертикальный перенос прокариотических симбионтов от материнского организма к дочернему, проникая в зародыш. Считается, что бактериоциты происходят из археоцитной линии .
Это гетерогенная группа клеток, содержащих в цитоплазме особые включения. В отличие от археоцитов, у них редуцирован шероховатый эндоплазматический ретикулум, нет фагосом, не выявлена активность кислой фосфатазы , в ядрах нет ядрышка. По размеру включений можно выделить клетки, содержащие крупные включения и клетки, содержащие мелкие включения. К первой группе относятся:
Ко второй группе относятся:
Способность большинства клеток, слагающих тело, к свободным перемещениям — отличительная черта губок. Клетки мезохила перемещаются почти постоянно. При перестройке водоносной системы свою локализацию могут менять эндопинакоциты и хоаноциты, благодаря чему образуются новые жгутиковые камеры, сливаются и образуются новые ветви каналов. Это обеспечивает оптимизацию тока воды в теле губки по мере её роста или в связи с изменениями в окружающей среде. Независимые и частые перемещения клеток в телах губок возможны благодаря отсутствию у большинства из них настоящих межклеточных контактов и базальной пластинки .

У стеклянных губок нет типичных хоанодермы и пинакодермы. Их живая ткань представляет собой трёхмерную сеть, похожую на паутину, которая образована многочисленными тяжами. Мембраны, разделяющие клетки, отсутствуют, и в пределах ткани цитоплазма непрерывна, поэтому ткань стеклянных губок называют трабекулярным синцитием или трабекулярной сетью. Вместо хоанодермы имеется ещё один синцитий — хоаносинцитий, а вместо типичных хоаноцитов — воротничковые тела, у каждого из которых есть воротничок и жгутик, но нет ядра. Воротничковые тела собраны группами в отдельных карманах, окружённых трабекулярной сетью. По сути, воротничковые тела — это выросты одной содержащей ядро стволовой клетки (хоанобласта). Мезохил тянется внутри каждого тяжа трабекулярной сети. Он содержит спикулы, коллагеновые волокна и отдельные клетки: склероциты, археоциты и, скорее всего, половые клетки .
Абсолютное большинство губок — это сидячие прикреплённые организмы, однако некоторые виды способны к ограниченной локомоции . Например, пресноводная губка Ephydatia , а также некоторые морские виды могут перемещаться по субстрату со скоростью от 1 до 4 мм в день. Такое перемещение достигается в результате амёбоидного движения пинакоцитов и клеток других типов, которое происходит в той или иной степени согласованно. Некоторые виды, например, Clathrina coriceae , могут сокращаться и изменять диаметр оскулюмов при помощи миоцитов. Благодаря этим механизмам губки могут временно приостанавливать ток воды через свою водоносную систему, например, при увеличении количества взвеси в воде . Взрослые губки всех четырёх классов могут изгонять из себя частички субстрата путём сокращения .
Показано, что изменения объёма и формы тела могут быть также эндогенными и иметь суточную ритмичность. Точно не известно, что отвечает за передачу сигнала между клетками в отсутствие нервной системы : клеточная проводимость, химическая диффузия или электрическая проводимость. По-видимому, любой из вариантов может иметь место, так как у губок выявлены биоэлектрические клеточные потенциалы и нейромедиаторы . Более того, в их геномах имеются гены , ортологичные генам, участвующим в функционировании нейросенсорной системы эуметазоев . Однако из-за отсутствия специализированных межклеточных контактов сигнал в теле губки распространяется очень медленно. Исключение составляют стеклянные губки, имеющие синцитиальное строение. Например, у Rhabdocalyptus dawsoni потенциалы действия быстро распространяются по синцитиальным тяжам, в результате чего останавливается биение жгутиков и ток воды через тело .
Большинство губок отфильтровывают мелкие пищевые частицы из воды, которая протекает через их тело. Размеры частиц обычно находятся в пределах от 50 до 1 мкм, но могут быть и ещё меньше. В эти пределы попадают одноклеточные планктонные организмы, например, динофлагелляты и бактерии, вирусы , мелкий органический детрит и, возможно, даже растворённые в воде органические соединения .
У губок нет популяции клеток, специализированной на выполнении пищеварительной функции. В захвате пищевых частиц участвуют почти все покровные клетки и многие клетки внутреннего пространства губки. Показано, что фагоцитарной и пищеварительной активностью обладают хоаноциты, археоциты, экзопинакоциты, эндопинакоциты и базопинакоциты. Захват пищи также может происходить в прозопилях и приводящих каналах . Фагоцитарная активность выявлена у миоцитов, секреторных глобулярных клеток, склероцитов, лофоцитов. Мелкие частички могут проникать в мезохил между экзопинакоцитами, однако механизм этого перемещения неясен. Некоторые клетки мезохила могут амёбоидно вытягивать свои отростки в полость приводящих каналов между пинакоцитами и осуществлять фагоцитоз. Однако в большинстве случаев бактерии и прочий пикопланктон захватываются псевдоподиальными выростами хоаноцитов .
Хоаноциты могут захватывать частички пищи и базальной частью, и апикальной (в области воротничка). Хоаноциты и пинакоциты, фагоцитировавшие пищевые частицы, или передают их археоцитам, или выделяют их в мезохил экзоцитозом . Археоциты и хоаноциты переваривают пищу в пищеварительных вакуолях , а археоциты, по-видимому, способны также запасать питательные вещества (гликоген и липиды ). Особо крупные частицы, оседающие в каналах, поглощаются эндопинакоцитами и археоцитами, которые проникают в каналы через слой пинакоцитов или через пороциты. В расщеплении пищи у губок принимают участие ферменты амилаза , липаза , протеаза , эстераза .
-pone.0035105.g003.jpg)
Среди губок известны и плотоядные виды — обыкновенные губки семейств Cladorhizidae , а также некоторые виды семейств Guitarridae и Esperiopsidae. Известно всего 137 видов хищных губок . Представители Cladorhizidae питаются рачками и другими мелкими животными, которые прилипают к длинным липким нитям клеточного строения, раскинутым губкой. Когда добыча приклеивается, нить укорачивается, подтягивая жертву к телу губки. Далее губка медленно обволакивает жертву и переваривает её, скорее всего, с помощью археоцитов. Нет никаких доказательств того, что губка использует яд или токсины . Некоторые хищные губки используют для ловли добычи крючкообразные спикулы . Хищные губки лишены хоаноцитов и водоносной системы , хотя губки рода используют свою видоизменённую водоносную систему для засасывания добычи внутрь тела. Большинство хищных губок — обитатели больших глубин (до 8840 м) .
Удалению из тела губки подлежат непереваренные остатки пищи, а также неорганические частицы, попавшие в губку с током воды. Последние удалять необходимо: они могут закупоривать приводящие каналы и нарушать их работу. Застрявшую в приводящем канале частицу фагоцитирует археоцит и далее переносит её ближе к выходу из водоносной системы, выделяя экзоцитозом в отводящий канал. У губок, использующих инородные частицы для постройки скелета, захваченная частица может быть перенесена к месту синтеза элементов скелета .
Поступление газов и продуктов обмена (главным образом, аммиака ) из тела в воду, протекающую по водоносной системе, обеспечивает простая диффузия . Она, возможно, играет некоторую роль и в распространении питательных веществ по телу губки, хотя в основном этим занимаются археоциты. У видов рода есть специальные внутренние волокна, по которым, как по рельсам, перемещаются археоциты, наполненные питательными веществами .
У стеклянных губок транспорт осуществляется в пределах синцития. После того, как пищевые частицы были захвачены воротничковыми телами и загружены в вакуоли, вакуоли перемещаются по всему синцитию вдоль микротрубочек с помощью моторного белка динеина . Таким же образом происходит перемещение везикул в псевдоподиях фораминифер и в аксонах нейронов некоторых высших животных .
Поскольку в пинакодерме и хоанодерме практически нет межклеточных контактов, эти слои клеток не формируют надёжный барьер, разграничивающий мезохил и воду; поэтому вода постоянно поступает внутрь тела губок. В клетках пресноводных губок обычно имеются сократительные вакуоли , осуществляющие осморегуляцию на уровне отдельных клеток, но не всего тела животного .
Губки обладают различными приспособлениями для регуляции тока воды через водоносную систему. У стеклянных губок жгутики перестают биться, если внутрь губки попадают частицы донных осадочных отложений или повреждена её ткань. При этом по синцитиальной ткани распространяется потенциал действия , и жгутики возобновляют биение тогда, когда их мембранный потенциал достигнет нормального значения. Возможно, потенциал действия вызывает вход кальция в воротничковые тела, и биение жгутика восстанавливается при откачивании кальция из клетки .
У губок же с клеточным строением тканей потенциал действия, насколько пока известно, не может распространяться по телу. Однако в ответ на плохое качество воды, физическое или химическое раздражение, или даже в спонтанном ритме такие губки могут медленно сокращать каналы своей водоносной системы, закрывать остии и оскулюм, в результате чего ток воды сильно замедляется или прекращается вовсе. При этом жгутики хоаноцитов не прекращают биений и работают непрестанно, хотя у некоторых хоаноцитов жгутики бьются медленнее, чем у других. Биение жгутиков происходит по синусоиде и в одной плоскости, причём в пределах одной хоаноцитной камеры жгутики каждого хоаноцита бьются в своей плоскости; однако в ответ на повреждающее воздействие (разрыв губки, нехватка кислорода) некоторые жгутики замедляют своё биение и могут даже остановиться на несколько часов. В настоящее время считается, что к мгновенной остановке биения жгутиков способны только стеклянные губки из-за своего синцитиального строения. Стеклянная губка Oopsacas minuta может очень медленно сокращать своё тело, причём сокращения вызываются волнами кальция, проходящими по синцитию. При сокращении тела губки хоаноцитные камеры и отдельные хоаноциты тоже сокращаются .
У губок имеется эффективная система врождённого иммунитета , которая позволяет губкам отличать прокариот- патогенов , прокариот-эндосимбионтов и прокариот, служащих пищей. Было показано, что при попадании в губку бактерии-эндосимбионты продуцируют особые вещества, например, в случае губки , которые подавляют иммунный ответ и апоптоз . В ответ на эти молекулы губка начинает обильно синтезировать белки эндоцитоза , вероятно, для контроля бактериальной популяции. Некоторые мембранные белки имеют эукариотические домены , ответственные за белок-белковые взаимодействия с эукариотическим внеклеточным матриксом и межклеточную адгезию , что, вероятно, играет роль в распознавании. Патогенные грамположительные и грамотрицательные бактерии несут (PAMP), благодаря которыми опознаются клетками губки. Например, липополисахариды (LPS), являющиеся PAMP грамотрицательных бактерий, распознаются у губок, как и у других Metazoa , Toll-подобными рецепторами , которые запускают иммунный ответ .
Губки отторгают трансплантаты от других видов, однако принимают трансплантаты от особей своего вида. У некоторых морских губок ключевую роль в отторжении инородного материала играют серые клетки. При внедрении чужеродного фрагмента серые клетки выделяют химические вещества, тормозящие перемещение клеток в поражённой области, чтобы клетки чужеродного фрагмента не могли воспользоваться транспортной системой губки. После этого серые клетки группируются и выделяют токсины, убивающие все клетки в поражённой области. Такое активированное состояние иммунной системы может сохраняться до трёх недель .
У губок есть как половое, так и бесполое размножение. Губки могут быть яйцеродящими и живородящими . Яйцеродящие губки обычно раздельнополы, а во втором случае нередко бывают гермафродитами . Впрочем, известен случай смены губкой её пола в течение года .
Гонад у губок нет, и образование гамет не приурочено к определённому участку мезохила, так что гаметы распределены по ней равномерно, но при этом всегда находятся от канала или камеры на расстоянии, покрываемом диффузией . Половые клетки образуются путём прямой трансформации из соматических клеток : ядрышковых амёбоцитов и хоаноцитов, но основные стадии развития гамет у губок сходны с таковыми у других животных .
При образовании мужских гамет ( сперматогенезе ) у стеклянных и обыкновенных губок, а также Homoscleromorpha образуются временные сферические образования, окружённые уплощёнными соматическими клетками — сперматоцисты, в которых и протекает сперматогенез. У известковых губок сперматоцист не формируется. В пределах сперматоцисты мужские половые клетки развиваются синхронно, но у Homoscleromorpha в пределах сперматоцисты наблюдается градиент созревания сперматозоидов, что также сближает эту группу губок с эуметазоями. У большинства губок сперматозоиды примитивного типа и имеют большой объём цитоплазмы с несколькими митохондриями ; акросомы , как правило, нет. Однако методами электронной микроскопии было установлено, что акросомы есть у сперматозоидов всех Homoscleromorpha и у некоторых обыкновенных губок. У известковых губок спермии атипичные, они имеют сферическую форму и лишены жгутика .
Развивающиеся женские гаметы располагаются в хоаносоме диффузно или мелкими группами. У большинства губок в оогенезе не происходит типичного образования желтка ; вместо него преобладают различные формы передачи ооцитам от вспомогательных клеток питательного материала, из которого образуется вторичный желток. При этом может происходить фагоцитоз целых клеток или их фрагментов, попадающих в фаголизосомы. Яйцо и вспомогательные клетки иногда заключены в особый фолликул, имеющий стенки клеточного строения . У Homoscleromorpha и яйцекладущих обыкновенных губок желток образуется за счёт эндогенного синтеза. В ооцитах ооплазма не сегрегирована. У яйцеживородящих губок нет яйцевых оболочек, но у яйцекладущих обыкновенных губок в ходе оогенеза образуется примитивная желточная оболочка .
В период размножения стенка сперматоцисты разрушается, и сперматозоиды попадают в отводящие каналы (реже — в атриум) и выбрасываются наружу через оскулюмы. Некоторые тропические губки внезапно извергают из оскулюмов молочные облачка спермы . Когда выброшенные сперматозоиды приближаются к другой губке, то вместе с токами воды они переносятся в водоносную систему. Внутри губки они подходят к поверхности хоанодермы или в жгутиковые камеры, где их фагоцитируют, но не переваривают, хоаноциты. Хоаноцит, поглотивший сперматозоид, теряет жгутик и воротничок и превращается в амёбоидную клетку (клетку-носительницу), которая переносит ядро сперматозоида к яйцеклетке и либо передаёт ей ядро, либо фагоцитируется яйцом . Некоторые губки выбрасывают яйцеклетки вместе со вспомогательными клетками в воду. Часто губки выбрасывают яйца и сперму в воду синхронизированно; это называют «курением» или «кипением» губок. У тропической рифовой губки синхронизация выброса спермы и яиц очень чёткая, так что происходящее раз в год и длящееся три дня «курение» можно предсказать заранее .
Бесполое размножение описано во всех классах губок и может осуществляться путём фрагментации, геммулогенеза и почкования .
Фрагментацию считают наиболее примитивным способом бесполого размножения. Она может происходить во время сильных штормов, из-за хищников (рыб, черепах), инфекционных заболеваний, разрыва или раскола субстрата, в результате чего тело губки распадается на фрагменты. Отделённые куски выживают благодаря способности губок к регенерации (подробнее см. ниже). Они прикрепляются к субстрату, перестраиваются и превращаются в новую губку . Поскольку фрагментация — это не закономерный этап развития губки, а событие, вызванное случайными внешними факторами, никаких цитологических изменений при фрагментации не происходит. Фрагментация наиболее характерна для обыкновенных губок .
Почкование — довольно редкий способ размножения губок: как облигатный он выступает в жизненном цикле лишь у представителей двух семейств ( и ), хотя на нерегулярной основе представлен во многих семействах . При этом у большинства губок (за исключением Homoscleromorpha и Radiospongilla cerebellata ) на теле материнской губки образуется плотное скопление клеток — в основном археоцитов. В основе формирования такой почки лежит миграция полипотентных клеток с их последующей дифференцировкой. Эта почка лишена хоаноцитных камер, каналов, оскулюма и после отделения от материнской губки прикрепляется к субстрату; там она начинает расти и формировать водоносную систему. При оседании почки могут сливаться друг с другом, в результате чего развиваются более крупные губки. У Clathrina почки формируются как вздутия концов трубочек, которыми представлено тело губки. У некоторых видов Tethya почки формируются на специальных стебельках, а у Oscarella и Aplysilla они представляют собой папиллы, которые отделяются от родительского организма и дают начало новым губкам . У Homoscleromorpha механизм формирования новой губки совершенно иной: у этих губок почки представляют собой выросты тела родительской губки, и при их образовании не происходит пролиферации , а клеточный состав почки идентичен клеточному составу родительской губки .

Геммулогенез — наиболее сложный тип бесполого размножения губок. Его результатом является образование геммул — покоящихся стадий, предназначенных для переживания неблагоприятных условий, а также распространения губок. Геммулы образуют пресноводные и некоторые морские губки в количестве сотен и тысяч . Снаружи геммула покрыта неклеточной защитной оболочкой, а внутри неё находится масса плюрипотентных клеток и тезоцитов (неактивных археоцитов), заполненных резервным веществом . Периодически тело губки отмирает и полностью распадаются, так что остаются только геммулы .
Геммулы чрезвычайно стойки ко многим неблагоприятным факторам и могут прорастать даже после двухмесячного содержания при температуре −80 °C и даже −100 °C. Четверть геммул прорастает после четырёхмесячного обезвоживания при температуре 5 °C . Специалисты, изучающие губок, нередко хранят увлажнённые геммулы в холодильнике и проращивают их для экспериментов или наблюдений .
Для многих видов, жизненный цикл которых включает бесполое размножение, характерно чередование полового и бесполого размножения. В частности, в пределах одной губки образование геммул начинается не раньше образования желтка в ооцитах из-за конкуренции за клеточный материал. Этим же обусловлено и снижение активности полового размножения во время бесполого размножения. У многих пресноводных губок бесполое размножение усиливается вслед за половым и завершает активную фазу жизненного цикла в популяциях губок, обитающих в нестабильных условиях .
По способности к регенерации губки превосходят всех прочих Metazoa. При повреждении тела губки большая часть археоцитов превращается в пинакоциты и хоаноциты, восстанавливая пинакодерму и хоанодерму; весь процесс занимает несколько дней. Способность к регенерации активно используется в аквакультуре туалетной губки, когда губку режут на части и уже через несколько месяцев из кусочков развиваются полноценные губки. Кроме того, высокая способность к регенерации губок используется исследователями, изучающими их физиологию. Кусочек губки кладут на покровное стекло, к которому губка прирастает, а потом накрывают часть губки вторым покровным стеклом. В итоге образуется очень тонкий слой живой материи, который удобно изучать под микроскопом. При исследовании отдельных клеток губки её расчленяют на отдельные клетки физическим или химическим путём. Если одну из таких клеток положить в сосуд, то на дне сосуда разовьётся одна молодая губка . Интересно, что клетки губок способны реагрегировать с образованием нормальной губки после разделения фрагмента губки на отдельные клетки. Промежуточной стадией этого процесса является образование примморфов — скоплений недифференцированных клеток, покрытых экзопинакодермой. Далее клетки претерпевают трансдифференировку с образованием нормальной губки, причём судьба клетки при трансдифференцировке определяется её положением в многоклеточном агрегате .
Яйцекладущие губки выводят зиготы во внешнюю среду, где и происходит их развитие. Для многих губок характерно живорождение, при котором зиготы остаются внутри материнского организма и покидают его уже сформированными личинками . У живородящих губок зародыши развиваются в особых выводковых камерах, сформированных уплощёнными клетками хоаноцитного или амёбоцитного происхождения, а также эндопинакоцитами. По окончании развития стенки выводковой камеры принимают участие в формировании каналов водоносной системы . Зародыши и личинки губок лецитотрофны, то есть питаются желтком . Губки демонстрируют широкое разнообразие вариантов эмбриогенеза и строения личинок, однако их развитие, несмотря на внешние различия, очень схоже на молекулярном уровне (паттерны экспрессии генов, перемещений и гибели клеток и т. п.) .
Зигота губок претерпевает полное и равномерное дробление, однако взаимное расположение бластомеров варьирует у разных представителей . В связи с этим у губок выделяют четыре основных способа дробления :
В связи со слабой специализацией яйцеклетки у всех губок (кроме Calcaronea) дробление, как правило, не характеризуется устойчивостью пространственного расположения бластомеров . Синхронность делений либо совсем отсутствует, либо рано утрачивается. На ранних этапах эмбриогенеза у многих губок из мезохила в зародыш проникают эндосимбиотические бактерии. В некоторых случаях в зародыш внедряются соматические клетки материнского происхождения .
У яйцекладущих обыкновенных губок родов Tetilla и Stylocordyla развитие прямое: после дробления сразу же наступает стадия дифференцировки клеток и развитие молодой губки .
При формировании многоклеточных зародышей (личинок) у губок описаны почти все типы клеточных перемещений, характерные для эуметазоев: морульная деламинация , клеточная деламинация, инвагинация и мультиполярная иммиграция. Однако у губок встречаются и уникальные морфогенезы, не встречающиеся у остальных многоклеточных животных — например, экскурвация, формирование псевдобластулы за счёт вселения материнских клеток в дробящийся зародыш, поляризованная деламинация, униполярная пролиферация и мультиполярная эмиграция .
Эмбриональное развитие губок принципиально отличается от такового у эуметазоев. В ходе эмбриогенеза губок не происходит формирования зародышевых листков , то есть отсутствует гаструляция и гаструла , хотя после дробления идёт интенсивный морфогенез и дифференцировка клеток . На поздних этапах дробления, которые обычно именуют бластулой или морулой, у губок начинается дифференциация личиночных клеток. Судьба специализированных личиночных клеток губок (покровных, скелетогенных и других) не гомологична судьбе зародышевых листков эуметазоев, потому что у губок нет частей тела, органов и тканей, гомологичных таковым у эуметазоев. Таким образом, у губок нет и не может быть эктодермы и энтодермы .
У губок описано восемь типов личинок: целобластула, кальцибластула, цинктобластула, амфибластула, дисферула, хоплитомелла, паренхимула и трихимелла .
Самые просто устроенные личинки — целобластула и кальцибластула, которые имеют однорядный жгутиковый покров и обширную внутреннюю полость (бластоцель), заполненную бесструктурной жидкостью, отдельными соматическими клетками материнского происхождения и симбиотическими бактериями. Пока целобластула ведёт планктонный образ жизни, некоторые жгутиковые клетки теряют жгутики и мигрируют в бластоцель, так что в конце концов полая целобластула превращается в лишённую полости . Целобластула описана у некоторых яйцекладущих обыкновенных губок, а кальцибластула присуща известковым губкам подкласса Calcinea. У кальцибластул в составе наружного слоя клеток могут находиться отдельные безжгутиковые клетки .
Другая однослойная личинка — амфибластула, она тоже представляет собой полый шар. Переднее полушарие амфибластулы сформировано маленькими жгутиковыми клетками, а заднее — безжгутиковыми крупными клетками с зернистой цитоплазмой. В области экватора расположены четыре клетки «креста». В небольшой внутренней полости находятся различные материнские клетки и симбиотические бактерии. Когда амфибластула находится в мезохиле родительской губки, все её жгутики обращены в бластоцель, однако вскоре образуется отверстие, через которое личинка выворачивается жгутиками наружу (инверсия). После этого личинка выходит наружу. Инверсия имеет место у тех губок, у которых яйца образовались из хоаноцитов . Амфибластулы свойственны известковым губкам подкласса Calcaronea .
Дисферула — одна из личинок группы обыкновенных губок Halisarca . Внутри личинки находится замкнутый слой жгутиковых клеток, который образовался в результате инвагинации (впячивания) поверхностного жгутикового покрова .
Хоплитомелла — это уникальная планктонная личинка сверлящих губок Alectona и . Она лишена покровных жгутиковых клеток. Внутри хоплитомеллы находятся хоаноцитные камеры, разнообразные клетки, волокна коллагена и спикулы типа дискотриенов, которых нет у взрослых губок .
Наиболее сложно устроены паренхимульные личинки, их организация может варьировать даже в пределах одного рода. Паренхимульные личинки характерны для подавляющего большинства обыкновенных губок. Паренхимула снаружи покрыта одно- или многорядным слоем одножгутиковых клеток, по строению соответствует гипотетической фагоцителле И. И. Мечникова . Внутри личинки располагаются склероциты, археоциты, секреторные клетки, колленциты, а также личиночный скелет, если он характерен для взрослой губки. В связи с этим паренхимулу можно в какой-то степени считать «почти готовой» молодой губкой . Паренхимулы, выходящие во внешнюю среду, характеризуются разной степенью дифференцировки личиночных клеток, а иногда и дефинитивных (то есть присущих взрослой губке) .
Трихимелла характерна для многих стеклянных губок . Принципиальные отличия трихимеллы от паренхимулы заключаются в наличии у трихимеллы в средней части пояса клеток, несущих до 50 ресничек . Вся личинка, в том числе и ресничные клетки, покрыта синцитиальным слоем пинакодермы, причём реснички выходят наружу через отверстия в синцитии. Внутри трихимеллы находятся жгутиковые камеры, образованные безъядерными воротничковыми телами, которые отличаются от таковых у взрослой губки, поскольку не связаны общей цитоплазмой. У трихимелл имеются спикулы стауроактины, отсутствующие у взрослых губок .
Для Homoscleromorpha характерна личинка цинктобластула, которая внешне похожа на целобластулу, поскольку образована одним слоем ресничных клеток и имеет обширную внутреннюю полость. Однако клетки наружного слоя соединены десмосомами в апикальной части, а со стороны полости подстилаются базальной пластинкой, поэтому их можно рассматривать как эпителиальные клетки. В районе заднего полюса личинки располагается пояс клеток с внутриядерными паракристаллическими телами. Внутренняя полость личинки содержит материнские клетки и симбиотические бактерии .
Для личинок всех типов, кроме хоплитомеллы, характерна переднезадняя полярность. Она проявляется в форме тела, характере ресничного покрова на разных полюсах, в неравномерности окраски, а также на гистологическом уровне (характер распределения различных типов клеток, ориентация и расположение спикул в случае паренхимул, неравномерном распределении симбиотических бактерий в полости личинки в случае цинкобластулы). Многие личиночные структуры при развитии личинки отмирают и не сохраняются у взрослых особей. К числу таких структур относится личиночный ресничный покров, выполняющий локомоторную, прикрепительную и, возможно, трофическую функции, а также личиночный скелет .
Все личинки губок — лецитотрофные: используют в качестве источника энергии гранулы желтка и липидные гранулы, не израсходованные в ходе эмбриогенеза. В связи с лецитотрофностью личинки губок живут относительно недолго . Впрочем, паренхимулы некоторых обыкновенных губок могут при помощи жгутиковых клеток фагоцитировать органические частицы из внешней среды . Обычно выход личинок стимулируется светом и происходит, как правило, на рассвете . Многие личинки движутся в воде штопорообразно. Особенности движения губок, скорее всего, обусловлены расположением ресничек, которое сильно варьирует у разных групп .
У личинки обыкновенной губки известно 3 типа клеток, которые могут выполнять сенсорную функцию. На заднем конце личинки имеются специализированные фоторецепторные клетки, образующие кольцо пигментированных клеток с фотосенсорными мембранами и длинными ресничками ; они регулируют фототаксис . Второй тип сенсорных клеток — глобулярные клетки, заполненные везикулами ; они экспрессируют ряд постсинаптических генов. Наконец, на переднем конце личинки располагается группа клеток, снабжённых одной ресничкой. Они соединены с другими клетками специальными контактами, но химических синапсов обнаружено не было .
Личинки губок могут быстро реагировать на стимулы из окружающей среды с помощью ряда ориентирующих поведенческих программ, таких как , фототаксис и реотаксис . Кроме того, они могут выбирать субстрат, на который впоследствии опустятся .
По прошествии определённого времени (от нескольких часов до нескольких дней) личинки оседают на дно и приступают к поискам подходящего места для прикрепления . Метаморфоз личинок губок начинается с прикрепления личинки к субстрату. Далее образуются экзопинакодерма и базопинакодерма, развиваются хоаноцитные камеры (или хоанодерма у асконоидных представителей) и каналы, соединяющиеся с ними, образуются поры и оскулюм. Одновременно происходит синтез спонгина, спикул и основного вещества мезохила. Однооскулюмная особь, формирующаяся непосредственно из личинки и имеющая водоносную систему, отличную от таковой у взрослых форм, у известковых губок имеет асконоидное строение и называется олинтус. Олинтус имеет единственную мешковидную хоаноцитную камеру и апикальный оскулюм. У обыкновенных губок и Homoscleromorpha эта стадия имеет сиконоидное или лейконоидное строение и называется рагон; он имеет один оскулюм и несколько хоаноцитных камер . У большинства паренхимульных личинок в ходе метаморфоза происходит деструкция покровных клеток, и новые покровы молодой губки формируются внутренними клетками. У амфибластул, кальцибластул, целобластул, цинктобластул и личинок Halisacra в ходе личиночного морфогенеза дифференцируются только наружные клетки .
Под типом развития понимается комплекс связанных друг с другом унаследованных и приобретённых морфогенетических процессов. У губок известно 8 типов развития, названных по характерным личинкам. Они перечислены в следующей таблице.
| Тип | Таксон | Яйцо | Дробление | Личинка | Метаморфоз | Бесполое размножение | Источник |
|---|---|---|---|---|---|---|---|
| Амфибластульный | Calcaronea (известковые губки) | Изолецитальное, олиголецитальное, овальной формы | Полное, асинхронное, неравномерное, инкурвационное | Амфибластула | По мезенхимному типу: жгутиковые клетки погружаются внутрь и дают начало хоаноцитам, клеткам мезохила, склероцитам и эндопинакоцитам, зернистые безжгутиковые клетки дают экзо- и базипинакодерму | Почкование | |
| Кальцибластульный | Calcinea (известковые губки) | Изолецитальное, олиголецитальное, без признаков поляризации, нет специальных питающих клеток | Полное, равномерное, полиаксиальное, имеется полость дробления | Исходно целобластульная личинка приобретает переде-заднюю полярность и становится кальцибластулой. Основной тип клеток личинки — жгутиковые клетки. Есть специальные межклеточные контакты. | По мезенхимному типу | Почкование | |
| Трихимелльный | Стеклянные губки | Изолецитальное, олиголецитальное, без признаков поляризации, нет специальных питающих клеток | Полное, равномерное, асинхронное | Трихимелла, имеются пробковые контакты | Не описан | Разные виды | |
| Дисферульный | Halisarcida (обыкновенные губки) | Изолецитальное, полилецитальное, без признаков поляризации, нет специальных питающих клеток | Полное, равномерное, асинхронное, полиаксиальное, имеется полость дробления. | Целобластула, паренхимула, дисферула. Личинки полностью покрыты жгутиками. Имеются адгезивные контакты | Смешанный: экзопинакодерма развивается по эпителиальному типу из жгутиковых клеток заднего полюса личинки, а базопинакодерма, экдопинакодерма и хоанодерма по мезенхимному | Не известно | |
| Паренхимульный | Обыкновенные губки | Строение варьирует | Полное, асинхронное, неупорядоченное | Паренхимула | По мезенхимному типу, сопровождается миграцией жгутиковых клеток внутрь и внутренних наружу. Полный или частичный фагоцитоз жгутиковых клеток личинки | Почкование, фрагментация, геммулогенез | |
| Прямое развитие | Обыкновенные губки | Мелкие, изолецитальные, олиголецитальные, без признаков поляризации | Полное, равномерное, асинхронное | Морульная деламинация | Нет | Почкование | |
| Целобластульный | Обыкновенные губки | Яйца окружены слоем коллагена и слизи, изолецитальные, олиголецитальные, неполяризованные | Полное, равномерное, может быть радиальным | Целобластула; адгезивные межклеточные контакты | Часть жгутиковых клеток выселяется внутрь и образует хоаноциты и пинакоциты, оставшиеся на поверхности — пинакоциты | Почкование | |
| Цинктобластульный | Homoscleromorpha | Изолецитальное, полилецитальное, полярность не выражена, специализированных питающих клеток нет | Полное, равномерное, асинхронное, неупорядоченное | Исходно целобластула, превращающаяся в цинктобластулу. Базальная пластинка, десмосомы | По эпителиальному типу | Почкование и фрагментация |








Продолжительность жизни губок колеблется у разных видов от нескольких недель и месяцев до десятков и сотен лет. В умеренных широтах она составляет от одного года до нескольких лет, причём пресноводные виды губок обычно живут всего несколько месяцев, но в некоторых случаях могут создавать многолетние образования особого рода. Однако морские тропические губки, а также глубоководные виды живут очень долго — до 200 лет и более . Рекордсменом долгожительства среди губок, вероятно, является — возраст экземпляра, добытого в 1986 году в Восточно-Китайском море на глубине 1110 м, оценивается в 11 (±3) тысяч лет .
Способность к половому размножению некоторые виды приобретают в возрасте 2—3 недель, а другие — только при достижении возраста в несколько лет. В целом губки растут относительно медленно (наибольшая скорость роста отмечена у форм с коротким сроком жизни). Рост и возраст губок сильно зависит также от внешних факторов — например, годичного перепада температур. Некоторые губки (Sclerospongia) отличаются очень низкой скоростью роста, давая прирост около 0,2 мм в год, так что возраст таких экземпляров диаметром 1 м может составлять 5000 лет . В то же время некоторые виды растут со скоростью в несколько дециметров за год. Молодая губка растёт сравнительно быстро, но с увеличением объёма тела скорость её роста уменьшается .
Особенности геномов губок были изучены на примере обыкновенных губок и . Они имеют относительно большие геномы (1670 мегабаз, масса геномной ДНК около 1,7 пг ). Кроме того, было показано, что геномы губок состоят в основном из последовательностей, представленных в геноме лишь по одному разу (без повторов) . Гены губок содержат мало интронов , особенно это касается генов, кодирующих белки , и их интроны довольно короткие: длина большинства из них составляет 0,1—2 килобазы, и ни один из найденных интронов не имел длину более 5 кб . Ядерный геном губок содержит гены , входящие в состав гомеобоксных генов. Ситуация с наличием у губок других ключевых для многоклеточных животных гомеобоксных генов — Hox -генов — менее ясна . У многих губок хромосомы очень маленькие, и в кариотипах обильно представлены микрохромосомы .
В митохондриальной ДНК (мтДНК) губок имеются интроны, хотя у большинства Metazoa интронов в мтДНК нет (исключения составляют только кораллы и актинии) . У известковых губок мтДНК имеет линейную структуру, а не кольцевую, как у большинства Metazoa. Так, у митохондриальная ДНК представлена 6 линейными хромосомами. Кроме того, у этой губки в митохондриях происходит модификация тРНК , а рРНК фрагментированы . Следует подчеркнуть, что у разных губок в митохондриях наблюдаются различные отличия от стандартного генетического кода . Так, у стеклянных губок кодон AGR кодирует не аргинин , а серин , а у известковых губок , Sycon coactum и Petrobiona massiliana — глицин или серин. Кроме того, у P. massiliana кодон AUA кодирует не изолейцин , а метионин . У известковых губок Clathrina clathrus и кодон UAG не терминаторный , а кодирует тирозин , а кодоны CGN (N — любой нуклеотид ) кодируют глицин вместо аргинина . Наконец, у известковой губки Leucosolenia complicata кодоны AGR начали кодировать метионин .
У губок идентифицированы многие гены и белки, ответственные за организацию их тела. Идентифицированы молекулы адгезии, обеспечивающие взаимодействие клеток друг с другом, а также с межклеточным матриксом . Важнейшая молекула, обеспечивающая взаимодействие двух клеток — фактор агрегации (AF). обеспечивает связь AF с мембранным клеточным рецептором агрегации (AR). Ключевые белки межклеточного матрикса — коллаген - и фибронектин -подобные белки — взаимодействуют интегринами , расположенными в клеточной мембране . У губок был идентифицирован по крайней мере один морфоген — , скорее всего, он участвует в формировании оси тела. Кроме того, у губок появляются многие семейства белков , характерные для прочих Metazoa: белки межклеточного матрикса (коллаген, белки, содержащие домены FN 3 -, SRCR, SCR); белки, участвующие в передаче сигнала ( тирозинкиназы ); белки врождённого иммунитета ( цитокины , (2—5)А-синтетаза, вариабельные -подобные домены) . Таким образом, данные молекулярной биологии указывают на то, что губки — это не неорганизованные скопления клеток, а сложно организованные животные с определённым планом тела .
У губок нет настоящих нейронов и нервной системы , однако у них имеются гены, связанные с функционированием нейронов, поведенческие реакции на внешние стимулы и сенсорные клетки, участвующие в восприятии сигналов и экспрессирующие гены, характерные для нейронов. обыкновенной губки , а также анализ транскриптомов губок различных классов показал, что у губок есть много генов, которые у стрекающих и двусторонне-симметричных животных характерны для клеток нервной системы. В их числе гены, связанные с быстрой синаптической передачей: потенциал-зависимые ионные каналы , нейросекреторные белки группы SNARE (синтаксин1, SNAP-25 , синаптобревин ); гены, связанные с первичным определением нейрогенных полей (в том числе белки ); ферменты , связанные с синтезом нейромедиаторов (в том числе декарбоксилаза ароматических аминокислот и триптофангидроксилаза ); большая часть генов постсинаптических окончаний ( NOS , цитрон и ) . При этом натриевые каналы , имеющиеся у хоанофлагеллят , у губок отсутствуют .

Губки распространены по всему миру. Большинство видов населяет океанские воды от полярных областей до тропических регионов . При этом наибольшего видового разнообразия губки достигают именно в тропических и субтропических районах Мирового океана . 219 видов (по другим данным, около 150 ) обитают в пресных водоёмах.
Большинство видов живёт в спокойных и чистых водах, поскольку частицы ила и песка, взвешенные в воде или поднятые со дна течениями, могут закупорить поры в телах губок, затруднив тем самым процессы дыхания и питания . Поскольку губки ведут прикреплённый сидячий образ жизни, для своего развития и роста они нуждаются в твёрдом субстрате. В связи с этим скопления губок возникают в местах, где на дне имеются каменистые материалы: камни, валуны, галька и т. п. Однако некоторые виды способны закрепляться в мягких осадочных придонных отложений при помощи корнеподобного основания тела .
Большинство губок обитают на небольших глубинах: до 100—500 м. С увеличением глубины количество видов губок уменьшается. На глубинах свыше 1000—1500 м губки обычно встречаются довольно редко, так что количество глубоководных видов невелико .
В умеренных водах губки более многочисленны, но менее разнообразны, чем в тропиках . Вероятно, это связано с тем, что в тропиках встречается немало организмов, питающихся губками . Стеклянные губки наиболее многочисленны в полярных водах, а также на больших глубинах умеренных и тропических морей, поскольку их пористое строение тела позволяет им с минимальными затратами извлекать пищевые частицы из этих бедных пищей вод. Обыкновенные губки и известковые губки многочисленны и разнообразны в более спокойных неполярных водах .
Губки, обитающие в приливо-отливных зонах, хорошо приспособлены к непродолжительному пребыванию на открытом воздухе, когда во время отлива они выступают из воды. При этом устья и поры у них закрываются, что предотвращает излишнюю потерю влаги и высыхание .
Среди эндемиков выделяются губки озера Байкал , представленные эндемичным семейством Lubomirskiidae , которое объединяет 4 рода и 13 видов .
Различия в местообитаниях различных классах губок указаны в следующей таблице :
| Тип вод | Глубина | Тип поверхности | |
|---|---|---|---|
| Известковые губки | Морские | Менее 100 м | Твёрдые |
| Стеклянные губки | Морские | Большие глубины | Мягкие или твёрдые осадки |
| Обыкновенные губки | Морские, пресные (около 80—90 % всех известных видов губок) | От приливной зоны до абиссали ; хищные обыкновенные губки могут быть найдены на глубине 8840 м | Любые |

Особым видом рифов являются губковые рифы . Подобные рифы ранее были широко распространены в истории Земли . Наиболее ранние из них датируются среднем триасом (245—208 млн лет назад), а своего наибольшего процветания они достигли в конце юрского периода (208—146 млн лет назад). В это время система губковых рифов, образованная преимущественно шестилучевыми губками и имеющая длину около 7 тысяч км, пересекала поперёк северный бассейн древнего океана Тетис ; она является наиболее крупной среди известных конструкций, созданных когда-либо живыми организмами. Резкое сокращение количества губковых рифов произошло в меловом периоде на фоне всё возрастающего доминирования коралловых рифов .
Мелководные губковые рифы ныне отсутствуют, и все известные подобные рифы являются глубоководными . Современные губковые рифы нуждаются в уникальных условиях существования, которые могут объяснить их редкость. Они возникают в желобах континентального шельфа , в местах с оптимальными для их развития условиями: со сравнительно низкой скоростью осадконакопления, с высокими скоростями течения , высокой концентрацией в воде диоксида кремния и низкой концентрацией кислорода, со стабильным морским дном, состоящим из камней, крупного гравия и крупных валунов, при температурном режиме воды 5,5-7,3 °C . Губковые рифы в настоящее время известны на западе Канадского континентального шельфа , широко распространены в Северной Атлантике, в канадской Арктике и на континентальном шельфе в Антарктике . Сформированный кремниевыми губками ( Demospongiae ) риф существует в Северном Ледовитом океане у острова Аксель-Хейберг . Четыре рифа, образованные шестилучевыми губками в заливе Королевы Шарлотты на расстоянии 70-80 км от береговой линии и на глубине 165—240 м, занимают площадь 700 км² морского дна . Некоторые губки из рода в северной части Тихого океана могут создавать рифы высотой более 20 м, простирающиеся на несколько километров. Также вдоль глубоководных коралловых рифов обычно тянется зона обломков кораллов , в которой доминируют губки, среди которых имеются виды, разрушающие мёртвые кораллы . В юго-западной части Баренцева моря имеются глубоководные сообщества губок и плеченогих .
Губки могут вступать в симбиотические отношения с представителями большинства групп организмов : с водорослями , грибами , прокариотами и др.
Представители типа Porifera предоставляют среду обитания для многих одноклеточных и многоклеточных организмов. Многоклеточные организмы обычно выступают комменсалами и поселяются на поверхности тела губки и в её внутренних полостях, не мешая её жизнедеятельности . Среди эндобионтов губок отмечены стрекающие , турбеллярии , немертины , сипункулиды , полихеты , моллюски , ракообразные , многоколенчатые , иглокожие , асцидии и рыбы .
Одноклеточными симбионтами губок могут быть динофлагелляты , зоохлореллы , красные водоросли , нитчатые зелёные водоросли , а также грибы . Пресноводные губки наиболее часто находятся в симбиотических отношениях с различными водорослями, которые обусловливают их зелёную окраску .
Губки, в которых поселяются фотосинтезирующие эндосимбионты, выделяют в три раза больше кислорода, чем поглощают, а также продуцируют больше органического вещества, чем потребляют. Такая роль губок в образовании первичной продукции особенно важна в австралийском Большом Барьерном рифе , но в Карибском море она не столь велика .
Однако особенно важными симбионтами губок являются облигатные видоспецифичные автотрофные или неавтотрофные прокариоты (бактерии и археи ), которые поселяются в мезохиле. Взаимодействие между губками и этими микроорганизмами может происходить по-разному. Уникальность симбиоза губок с прокариотами заключается в том, что все исследованные к настоящему времени виды губок имеют симбиотические ассоциации с одним или более видами бактериальных симбионтов .
Прокариотические симбионты, как правило, располагаются между элементами мезохила и физически отделены от воды пинакодермой. Однако известны губки, у которых симбионты внутриклеточные: они локализованы в специальных клетках — бактериоцитах . Биомасса прокариотических эндосимбионтов может быть весьма значительной и составлять до 40 % (и даже до 60 %) массы губки-хозяина, а концентрация бактериальных симбионтов в губке может на 2-3 порядка превышать концентрацию прокариотических организмов в окружающей водной среде . Прокариоты могут проникать в тело губки двумя путями: из воды в окружающей среде, или же путём вертикального переноса от материнской губки к следующему поколению через яйца или личинки; возможность вертикального переноса прокариотических симбионтов говорит об их исключительной важности для жизни губок. Губкам удаётся поддерживать разнообразные, но в то же время специфичные микробные сообщества внутри себя — несмотря на то, что вместе с током воды при фильтрационном питании в их тела проникает множество морских микроорганизмов .
Внутри губок поселяются прокариоты самого разнообразного систематического положения, относящиеся примерно к 20 бактериальным филам , причём представители одной филы — Poribacteria — встречаются исключительно в губках. Симбиотические прокариоты могут принимать участие в разнообразных физиологических процессах губок — например, в круговороте нерастворимых белков и структурных перестройках органического скелета и внеклеточного матрикса. Многие прокариоты-симбиоты синтезируют антибиотики и вещества с резким запахом, отпугивающим хищников. Они также могут служить дополнительным источником питания губок, причём могут поглощаться как сами прокариоты путём фагоцитоза, так и продукты их жизнедеятельности . Можно сказать, что губка-хозяин и населяющие её микроорганизмы метаболически связаны и обмениваются соединениями азота , продуктами фиксации СО 2 , вторичными метаболитами и полученными из воды органическими соединениями . Например, внутри губки Lophophysema eversa живут автотрофные микроорганизмы, способные окислять аммоний , нитрит и сульфид и потому играющие важную роль в круговороте углерода, азота и серы в теле губки. Кроме того, они нейтрализуют токсичные для губки-хозяина аммоний, нитрит и сульфид .
Губки вступают в разнообразные взаимоотношения с другими животными. Например, довольно крупная губка , живущая на каменистых субстратах, может также заселять заросли морских трав . При этом она позволяет окружать и обрастать себя губкам, живущим в морских травах и опасным для местных морских звёзд ; таким образом, Lissodendoryx colombiensis обеспечивает себе защиту, а губки зарослей морских трав получают возможность жить на некотором поднятии относительно морского дна .

Особенным разнообразием отличаются взаимоотношения между ракообразными и губками . Многие виды мелких веслоногих и равноногих рачков, бокоплавов сожительствуют или паразитируют в губках. Часто десятиногие раки и креветки используют губок в качестве убежища от врагов. Креветки рода Synalpheus из семейства Alpheidae образуют колонии внутри губок, причём каждый вид креветок населяет только определённый вид губок; благодаря этому род Synalpheus является одним из самых многочисленных родов ракообразных . Вид Synalpheus regalis использует губок не только для питания, но также для защиты от других креветок и хищников. В одной губке может жить 16 тысяч особей креветок, питающихся самыми крупными частицами, попадающими внутрь тела губки при фильтрационном питании . В другом семействе креветок — Spongicolidae — практически все представители являются облигатными симбионтами глубоководных стеклянных губок (только в роде известны свободноживущие формы) . В частности, самец и самка из рода поселяются в полости стеклянной губки на стадии личинки, а во взрослом состоянии выйти за остов губки уже не могут .
_-_NHM_London_-_With_sponge.jpg)
Непривлекательность губок для хищников часто используют некоторые крабы (например, Dromia personata ), которые носят на себе куски губок, поддерживая их задней парой своих конечностей. Со временем губка приобретает форму панциря . Острорылый краб прикрепляет губок, водоросли и сидячих животных к своему панцирю, так что на его спине образуется микросообщество, маскирующее краба . 13 видов раков-отшельников могут образовывать симбиоз в форме комменсализма с пробковой губкой ( ) . Последняя селится на пустой раковине брюхоногого моллюска, и в этой же раковине поселяется рак-отшельник. Со временем вся раковина обрастает губкой; внутри последней образуется спиральная полость, в которой живёт рак-отшельник .
Некоторые животные используют в качестве защиты отдельные иглы губок, в массе откладывающиеся после их гибели на дне. Эти животные сооружают свои домики частично или полностью из спикул и игл губок ( фораминиферы , некоторые черви, голотурия и др.) .
Иногда на губках могут поселяться сидячие, прикреплённые животные, к числу которых относятся различные усоногие раки (в частности, балянусы ). На корневом пучке игл ряда видов стеклянных губок часто встречаются морские уточки , а также могут селиться (Palythoa). На некоторых губках ( ) постоянно поселяется корковая актиния ( ) .

.jpg)

Многие губки имеют спикулы, которые защищают их от поедания иглокожими и другими хищниками . Тем не менее, морские черепахи охотно питаются губками, и кремнезёмные спикулы могут составлять до 95 % их фекалий. Губки являются типичными пассивно-ядовитыми животными, использующими для защиты от врагов свои токсичные метаболиты , включая природные . Первым токсином, выделенным из губок ( ) стало вещество, получившее название . Токсины группы бромфакелинов впервые были выделены из губки ; токсин, близкий по структуре обнаружен в губке . Пиррольные алкалоиды широко встречаются среди губок, особенно представителей семейства . Показано, что 9 из 16 антарктических губок и 27 из 36 карибских губок токсичны для рыб . В то же время среди рыб встречаются виды, специализированные на питании губками ( мавританский идол , рыбы-ангелы рода , рыбы-бабочки рода и др.) . Голожаберные моллюски группы проявляют узкую специализацию, питаясь только определёнными видами губок.
Токсины, выделяемые губками, могут быть направлены также против других сидячих организмов (например, мшанок и асцидий ), чтобы они не поселялись на губках и в непосредственной близости от них, благодаря чему губки могут успешно завоёвывать новые площади для жизни. Примерами таких токсинов могут служить и дибромагеласпонгин, синтезируемые губками из рода . Карибская губка выделяет токсины, которые убивают коралловые полипы , и губка растёт на их скелетах. Органические соединения, похожие по своей структуре на , а также 2-бромалдизин, характеризующиеся умеренным противомикробным действием, найдены в различных видах губок .
Многие губки обладают сильным запахом, например, «чесночная» губка . Некоторые виды могут вызывать у человека дерматиты при непосредственном контакте . Например, прикосновение к карибской «огненной» губке может вызвать сильное раздражение кожи .
Стеклянные губки не синтезируют токсинов, потому что на больших глубинах, в которых они живут, хищников мало .

Существует семейство сетчатокрылых насекомых — сизириды (Sisyridae), представители которого паразитируют на пресноводных губках. Самка откладывает яйца на растения , свисающие над водой. После вылупления личинки попадают в воду и ищут губок, которыми будут питаться. Их удлинённый ротовой аппарат предназначен для того, чтобы пробивать покровы губки и сосать жидкость из её тела. У некоторых видов личинки прикрепляются к поверхности губки, в то время как у других они поселяются во внутренних полостях её тела. Полностью сформированная личинка выходит из воды и свивает кокон, в котором окукливается .
Преимущественно губками питаются брюхоногие моллюски из семейства плевротомарий . Специализируются на питании губками и некоторые морские звёзды (например, виды рода ) .
Наиболее часто заболевания отмечаются среди губок Папуа-Новой Гвинеи , Мексики и Большого Барьерного рифа . Несмотря на огромное экологическое и экономическое значение, заболевания губок изучены плохо. Известно, что в роли патогенов могут выступать грибы , вирусы , бактерии (включая штаммы родов Bacillus и ). Показана связь эпидемий заболеваний среди губок и условий окружающей среды, таких как изменение климата .
Карибские губки рода страдают от особого заболевания, известного как «синдром красных полос». На телах больных губок образуется одна или несколько полос ржавого цвета, иногда окружённые полосами мёртвых тканей. Тело губки может быть целиком окружено такими поражениями. Болезнь, по-видимому, является заразной, и на Багамских рифах заражены около 10 % особей . Яркий цвет полосы имеют из-за цианобактерии , однако неизвестно, ответственна ли она за возникновение болезни .
Сверлящие губки семейства секретируют особые вещества, продуцируемые археоцитами и позволяющие им вбуравливаться в камни, кораллы и раковины мёртвых моллюсков .
Дельфины афалины из залива Шарк в Западной Австралии используют губки интересным образом: они прикрепляют губки к своему роструму, чтобы защитить его от повреждений во время поиска пищи в песчаном дне . Этот вид поведения, получивший названия спонгинг, известен только у афалин из залива Шарк, причём чаще всего спонгинг демонстрируется самками. Было показано, что матери учат спонгингу своих дочерей, причём все дельфины, демонстрирующие спонгинг, приходятся друг другу близкими родственниками (так что спонгинг, по-видимому, является недавним приобретением) .

Известно, что обыкновенные губки, но не эуметазои, могут синтезировать — стабильное производное 24-изопропил холестерола . Было показано, что к синтезу 24-изопропилхолестана неспособны и хоанофлагелляты — ближайшие одноклеточные родственники животных, поэтому по наличию 24-изопропилхолестана в ископаемых остатках можно судить о существовании губок в то время . По данным 2005 года, древнейшие находки 24-изопропилхолестана имеют возраст 1800 млн лет , однако более точный анализ показал, что этот биомаркер появился лишь незадолго до окончания протерозойского оледенения , то есть 635 млн лет назад . Таким образом, доказательства существования губок до протерозойского оледенения отсутствуют. Более того, 24-изопропилхолестан — «биомаркер губок» — мог синтезироваться и другими организмами, например, морскими водорослями .
Хотя молекулярные часы и биомаркеры свидетельствуют о том, что губки существовали задолго до кембрийского взрыва , кремниевые спикулы, похожие на спикулы обыкновенных губок, появляются в геологической летописи только начиная с кембрия . Впрочем, имеются неподтверждённые данные о находках кремниевых спикул в камнях, имеющих возраст 750 миллионов лет . Хорошо сохранившиеся древнейшие ископаемые остатки губок, найденные в формации Доушаньто , имеют возраст 580 млн лет и относятся к эдиакарскому периоду . Эти ископаемые остатки, в которых различимы спикулы, пинакоциты, пороциты, археоциты, склероциты и внутренние полости, были отнесены к обыкновенным губкам. Другие возможные обыкновенные губки были найдены в раннекембрийских отложениях в маотяньшаньских сланцах и имели возраст от 525 до 520 млн лет . Древнейшие ископаемые остатки стеклянных губок имеют возраст 540 млн лет и были найдены в каменистых породах Австралии, Китая и Монголии . У ранних кембрийских губок из Мексики , относящихся к роду Kiwetinokia , подтверждено слияние нескольких маленьких спикул с образованием одной большой спикулы . Спикулы из карбоната кальция, присущие известковым губкам, были найдены в Австралии в ранних кембрийских породах возрастом от 530 до 523 млн лет. Пресноводные губки появились гораздо позже. Самые ранние ископаемые остатки пресноводных губок относятся к среднему эоцену и имеют возраст от 48 до 40 млн лет . Хотя 90 % современных губок относятся к обыкновенным губкам, ископаемые остатки представителей этой группы более редки, чем остатки губок других групп, поскольку их скелет образован относительно мягким, плохо фоссилизирующимся спонгином . Появление эндосимбионтов внутри губок относится к раннему силуру .
Есть признаки, того, что губки могли существовать уже 890 млн лет назад, однако этот вопрос требует дополнительных исследований .

|
Современные представления о месте Porifera в системе Metazoa
|
|||||||||||||||||||||||||||||||||||||||
|
Как уже отмечалось в разделе «История изучения», на протяжении бо́льшей части XX века губок (тип Porifera) включали в состав Metazoa (настоящих животных) и противопоставляли в качестве группы Parazoa всем остальным их представителям ( Eumetazoa ) . При этом монофилия Porifera обычно не ставилась под сомнение , а вот в отношении Metazoa в целом такие сомнения возникали. В 1970—1980 гг. ряд зоологов придерживался гипотезы о независимом происхождении губок (совместно с хоанофлагеллятами ) и Eumetazoa от одноклеточных предков (предположительно, от ) .
Применение методов молекулярной филогенетики положило конец сомнениям в единстве Metazoa; выяснилось также, что хоанофлагелляты — сестринская группа для Metazoa в целом . Однако данные молекулярных исследований в отношении монофилии губок оказались противоречивыми. Так, результаты ряда исследований, полученных начиная с 2001 года, свидетельствовали о том, что одни группы губок в систематическом отношении ближе к эуметазоям, чем другие группы. Это означало парафилию Porifera: получалось, что последний общий предок губок был предком не только губок, но и эуметазоев. Анализ генов рРНК, проведённый в 2001 году, показал, что наиболее фундаментальные отличия внутри губок имеются между стеклянными губками и всеми остальными губками, а эуметазои наиболее близки к известковым губкам . Два анализа 2007 года, основанных на отличиях в генах рРНК и в спикулах соответственно, показали, что стеклянные губки и обыкновенные губки гораздо более близки друг к другу, чем каждая из этих групп к известковым губкам, а последние, в свою очередь, наиболее близки к эуметазоям .
Другие анатомические и биохимические данные говорят о близости эуметазоев и Homoscleromorpha — класса губок, который долгое время рассматривали в составе обыкновенных губок. Анализ ядерной ДНК, проведённый в 2007 году и не включающий стеклянных губок и гребневиков , показал, что самой близкой к эуметазоям группой губок является класс Homoscleromorpha, следующей по близости — известковые губки, а обыкновенные губки являются эволюционными «тётками» для обеих групп . Сперматозоиды Homoscleromorpha имеют ряд общих черт со сперматозоидами эуметазоев, которые отсутствуют у всех прочих губок. И у Homoscleromorpha, и у эуметазоев слои клеток прикрепляются к похожей на ковёр базальной мембране, состоящей в основном из белка коллагена типа IV, который не обнаружен у остальных губок. Впрочем, спонгин, поддерживающий мезохил обыкновенных губок, близок к коллагену типа IV .
Итак, согласно приведённым выше данным, губки представляют собой наиболее ранние ответвления клады настоящих многоклеточных животных (Metazoa). Однако в 2008 году был проведён анализ 150 генов 21 биологического вида (от грибов до человека; губки в этом анализе были представлены всего двумя видами), который показал, что в самом основании древа Metazoa лежат не губки, а гребневики. Если его данные верны, то либо гребневики получили все свои сложные структуры независимо от Metazoa, либо предки губок имели гораздо более сложное строение, и все современные губки представляют собой кардинально упрощённые формы . Полученные позднее данные поддерживают версию о том, что гребневики являются группой, сестринской к остальным метазоям . Последние данные по морфологии, особенностям развития и молекулярной биологии (в частности, анализ митохондриальной рибосомной ДНК, а также 18S и 28S ядерной митохондриальной ДНК ) говорят о том, что губки всё же являются монофилетической группой и вместе со стрекающими выступают как сестринские группы по отношению к двустороне-симметричным животным .
В пределах типа губок стеклянные и обыкновенные губки считаются наиболее близкими друг к другу как по молекулярным, так и по анатомическим и палеонтологическим данным. Подтверждена монофилия стеклянных губок, известковых губок и двух подклассов известковых губок (Calcinea и Calcaronea). Однако филогенетические отношения в пределах обыкновенных губок подверглись существенным изменениям. Чётко монофилетическая группа Homoscleromorpha была выделена из обыкновенных губок в ранге класса, а оставшиеся в составе Demospongiae отряды и семейства также образуют монофилетическую группу, причём они подразделяются на четыре кластера уровнем выше отряда , рассматриваемые сейчас в ранге подклассов . Что же касается типа Porifera в целом, то имеющиеся данные не позволяют сделать однозначный вывод, является ли данный таксон монофилетическим или парафилетическим ; в тех исследованиях, которые свидетельствуют в пользу монофилии губок, Homoscleromorpha выступают как сестринская группа по отношению к известковым губкам. В любом случае все четыре выделяемых ныне класса губок дивергировали друг от друга свыше 600 млн лет тому назад .
Археоциаты — это группа животных, чьи ископаемые остатки весьма многочисленны в отложениях раннего кембрия (от 530 до 520 миллионов лет назад), но полностью отсутствуют в отложениях позднего кембрия. Им приписывали родство с губками, стрекающими, водорослями и фораминиферами и предлагали выделить в отдельный тип (Archaeocyatha) животных или даже царство (Archaeata или Inferibionta). Однако с 1990-х годов археоциатов рассматривают как своеобразную ископаемую группу губок .
Ранее к губкам относили группу кембрийских мешковидных организмов, известных как . На основании сходства склеритов и спонгиновых волокон обыкновенных губок эти две группы сближали , однако в 2002 году появились данные о том, что Chancelloriidae нельзя рассматривать в составе губок, потому что эти организмы являются промежуточным звеном между губками и более высокоорганизованными животными. В качестве одного из доказательств последнего положения приводилось строение покровов Chancelloriidae: они более толстые и плотные, чем у губок . В 2008 году был проведён детальный анализ склеритов Chancelloriidae, который показал, что они очень близки по строению к склеритам — группы подвижных двусторонне-симметричных животных, похожих по виду на звенья кольчуги и известных по ископаемым остаткам из раннего и среднего кембрия .

Карл Линней, относивший большинство сидячих животных к порядку Zoophyta класса Vermes, ошибочно поместил род обыкновенных губок к растениям в порядок Algae (водоросли) . Впоследствии долгое время губок выделяли в отдельное подцарство Parazoa царства животных, противопоставляя их эуметазоям, к которым относятся все остальные животные (кроме пластинчатых ) .
В настоящее время известно около 8000 (по другим оценкам, более 15000 ) видов губок, из которых 219 (по другим данным, 150 ) — пресноводные. Современные виды губок разделены на четыре класса .
-pone.0035105.g007.jpg)
В состав класса известковые губки ( лат. Calcarea ) входит 680 видов, объединённых в 5 отрядов ( , , , Clathrinida , Murrayonida ), 24 семейства и 577 родов, что составляет примерно 8,2 % от всех описанных видов губок. Класс признан монофилетическим. Известковые губки — исключительно морские организмы, населяют все акватории от литорали до батиали . Минеральный скелет данных губок состоит только из карбоната кальция. Спикулы (двух-, трёх- и четырёхлучевые) располагаются в мезохиле свободно; иногда спаянные друг с другом спикулы формируют плотный скелет. В классе представлены все варианты строения водоносной системы. Личинка однослойная, с полостью внутри (амфибластула или кальцибластула). Класс подразделяется на два монофилетических подкласса — Calcaronea (личинка амфибластула; первые три перечисленных выше отряда) и Calcinea (личинка кальцибластула; два оставшихся отряда) .

Губки класса Homoscleromorpha раньше причислялись в составе подкласса к обыкновенным губкам, однако современные филогенетические исследования показывают, что это — чётко ограниченная группа губок, независимая от обыкновенных губок. К данному классу относится один отряд Homosclerophorida с двумя семействами и 6 родами. Представители класса — небольшие гладкие губки с формой тела от корковой до комковидной, обитающие в морях на небольших глубинах. Водоносная система — силлеибидная или лейконоидная. Может иметься неорганический кремнезёмный скелет, представленный мелкими четырёхостными спикулами и их производными, причём лучи спикул могут ветвиться и изгибаться. Базальная часть клеток хоанодермы и пинакодермы подстилается базальной пластинкой, сами клетки соединяются специализированными контактами. Базальная пластинка содержит коллаген типа IV, тенасцин и ламинин. Такое строение хоанодермы и пинакодермы соответствует строению эпителия эуметазоев. В период размножения базальная пластинка покрывает сперматоцисту и имеется у личинок. Экзо- и эндопинакоциты снабжены жгутиками. Характерно незначительное разнообразие клеточных типов. Входящие в класс губки являются живородящими, личинка цинктобластула. Бесполое размножение — в форме почкования и фрагментации .
-pone.0035105.g002.jpg)
К классу обыкновенные губки ( лат. Demospongiae ) относится 83,3 % всех современных губок — около 6900 видов (объединяются в 12 отрядов: Agelasida , Astrophorida , , Dendroceratida , Dictyoceratida , , , Haplosclerida , , Poecilosclerida , , Verongida ). Морские и пресноводные обитатели. Скелет только спонгиновый или спонгиновый с кремниевыми спикулами, которые в зависимости от размера подразделяют на макросклеры и микросклеры. Спонгин формирует отдельные волокна или объединяет спикулы. Некоторые формы полностью лишены скелетных элементов. Форма тела разнообразна; водоносная система в основном — лейконоидная, однако хищные губки из отряда Poecilosclerida полностью утратили водоносную систему. Личинки в основном паренхимулы, но у некоторых групп имеются однослойные личинки. У разных форм встречаются живорождение, яйцеживорождение и наружное развитие. В составе Demospongiae выделяют четыре подкласса: (отряды Dendroceratida и Dictyoceratida), Myxospongiae (или : отряды Chondrosida и Verongida), Haploscleromorpha (отряд Haplosclerida) и (остальные отряды обыкновенных губок) .
-pone.0035105.g005.jpg)
В состав класса стеклянные губки ( лат. Hexactinellida, Hyalospongia ) входит около 600 видов (7,3 % от всех описанных видов Porifera), собранных в 6 отрядов ( , , , , , ), 26 семейств и 166 родов. Обитают в морях на глубинах от 5 до 6770 м. Скелетные элементы представлены в основном шестилучевыми спикулами и их производными. Макросклеры, спаянные друг с другом, формируют жёсткие скелетные структуры. Большинство клеток объединяются в синцитий, а отдельные клетки, располагающиеся в карманах или капсулах синцития, могут быть представлены особыми поровыми контактами. Водоносная система сиконоидная, силлеибидная или лейконоидная. Все представители класса живородящие, личинка трихимелла. Выделяют 2 подкласса стеклянных губок: и . Названия подклассов связаны с формой микросклер: у представителей подкласса Amphidiscophora (содержит первый из перечисленных отрядов) микросклеры представляют собой амфидиски (тонкие стержни, на обоих концах которых расположены венчики из нескольких лопастей), а у губок подкласса Hexasterophora (5 оставшихся отрядов) — гексастры (мелкие шестилучевые иголочки, у которых лучи нередко несут на концах разнообразные придатки) .

Целый ряд факторов может негативно влиять на размер популяций губок, вызывать их угнетение и гибель. В настоящее время главной угрозой сокращения численности многих видов является хозяйственная деятельность человека. В частности, угрозу для губок представляет траловый промысел , при котором уловы губок могут доходить до 5—12,5 т на морскую милю . Под его воздействием в середине XX века всего за нескольких десятилетий во многих местах мирового океана произошла резкая деградация поселений губок; известны многочисленные факты полного разрушения губковых биотопов . В результате данного вида промысла в Баренцевом море суммарная биомасса губок (преимущественно и ) сократилась на два порядка , а в центральной части моря массовые поселения губок практически полностью исчезли .
В 2002 году был принят добровольный и регулируемый запрет донного траления в районах, где известны глубоководные губковые рифы. Тем не менее, добровольный отказ рыбаков не является полностью эффективным методом .
Опасность для губок представляет и загрязнение мирового океана , особенно прибрежных вод в густонаселённых регионах. Так, повышение эвтрофикации вследствие загрязнения сточными водами прибрежных мелководий Чёрного моря и последовавшее снижение концентрации в воде кислорода привели к снижению численности в этом регионе широко распространённой в Арктике и Северной Атлантике губки Halichondria panicea ( морской каравай ), которая в настоящее время как уязвимый вид занесена в .

Спикулы из карбоната кальция или кремнезёма делают большинство губок слишком жёсткими для какого-либо использования, однако у двух губок — и туалетной губки ( Spongia ) — скелеты мягкие. В Европе эти губки издревле нашли разнообразное применение. Их использовали в качестве подкладки под шлем, переносного сосуда для питья, а также в качестве фильтров городской воды. До изобретения синтетических губок они применялись для мытья, в качестве средства для нанесения краски и глазури . В зоотехнии и ветеринарии губки применялись для сбора спермы при искусственном оплодотворении скота . Сухие растёртые губки служили абразивным материалом для шлифовки металлических, деревянных и глиняных изделий . Однако из-за чрезмерного промысла к середине XX века губки и связанная с ними индустрия оказалась на грани исчезновения .
С давних времён губки нашли применение в медицине. Самые мягкие и тонкие виды рода применялись в хирургии для остановки кровотечений , а также в качестве средства контрацепции для предотвращения беременности . Ныне губки сохраняют важное медицинское значение, поскольку и сами они, и населяющие их эндосимбионты синтезируют множество биологически активных соединений, обладающих противовирусными, антибактериальными, противогрибковыми и даже противоопухолевыми свойствами . Возможно, необходимость синтеза таких необычных соединений заключается в отсутствии у губок надёжной защиты. Морские губки являются самым богатым источником фармакологически активных соединений среди всех морских организмов. На 2010 год из самих губок или их симбионтов было выделено более 5300 различных веществ, и ежегодно этот список пополняется более чем 200 наименованиями .

Примером синтезируемых губками соединений могут служить окисленные производные жирных кислот — , которые обладают противораковой, антибактериальной и противогрибковой активностью. , соединение группы оксилипинов, выделенное из губок, оказалось токсичным для клеток лимфомы у мышей . Губка синтезирует , синтетический аналог которого — — обладает противораковыми свойствами . Показано, что некоторые алкалоиды , выделенные из губок, помогают защитить почки от цитотоксичного действия противоракового препарата цисплатина . Алкалоиды губки рода ( B, C, D) убивают покоящиеся формы возбудителя туберкулёза Mycobacterium tuberculosis . Такой же активностью обладает неамфамид B — циклический губки . При этом , синтезируемый губкой , подавляет ВИЧ . Губка синтезирует галистанолтрисульфат — антибиотик широкого спектра действия, разрушающий цитоплазматическую мембрану . Губка образует вещества с антибактериальным и антигрибковым действием . Спонготимидин и спонгоуридин, выделенный из стали прообразом для синтезирования противовирусных средств видарабин , цитарабин и зидовудин . Более 100 веществ, выделенных из губок и имеющих противоопухолевый эффект, а также их синтетические аналоги и производные исследованы in vitro на различных линиях опухолевых клеток . Более 60 вторичных метаболитов губок обладают противомалярийной активностью : например, Аксизонитрил-1 из губки стал родоначальником изонитрилсодержащих производных препаратов и их аналогов .
Микроорганизмы, живущие внутри губок, могут служить природными резервуарами генов устойчивости к антибиотикам , как в случае губки .
В настоящее время проходят исследования перспективности использования хитинового скелета морских губок семейства при создании биоинженерных конструкций жировой , хрящевой и костной тканей на основе мезенхимных стромальных клеток .
Растёртые в порошок высушенные пресноводные губки бадяги — традиционное народное средство от синяков , ушибов , ревматизма , которое применялись для растирания кожи, либо как мазь в смеси с жиром или водой. Лекарство из бадяги использовалось также как внутреннее средство для лечения золотухи . В настоящее время промышленно изготавливается одноимённый лекарственный препарат , обладающий местным раздражающим, противовоспалительным и анальгезирующим действием. Порошок из сушёных бадяг, несмотря на вред при длительном употреблении, также традиционно использовался в качестве косметического средства и дешёвой замены румян . Гончары Южной Америки добавляли этот порошок в гончарную глину для повышения прочности изделий из неё .
Начиная с 1907 года губки использовались исследователями для изучения межклеточной адгезии, а также адгезии клеток и межклеточного матрикса . Губки могут служить удобными объектами для исследователей, в качестве модели для изучения более сложных животных . Благодаря своей пористой структуре, похожей на структуру костной ткани , наличию спонгина — очень похожего на коллаген животных полимера, широко применяющегося при регенерации тканей, и минерального компонента, губки хорошо подходят для использования в экспериментах по созданию костной ткани .
В культуру и быт народов Средиземноморья использование морских губок прочно вошло задолго до начала нашей эры — прежде всего, в качестве средства личной гигиены : при купании и нанесении на тело оливкового масла и духов (в данном качестве используется освобождённый от остальных частей тела губки её скелет, который состоит из сети роговых волокон и напоминает эластичный войлок ). Древние египтяне применяли губку при бальзамировании покойных, натирая тела ароматическими маслами. Гомер упоминает губки в обоих своих поэмах: в « Илиаде » губку дают в руки Гефесту , который с её помощью очищает свои руки, лицо и грудь, а в « Одиссее » слуги Одиссея используют её для очистки столов в его доме после визита к Пенелопе женихов. Римские воины подкладывали губки под свои шлемы , а врачи применяли их для очистки ран .
Кроме этого, губки в античные времена считались одним из инструментов, применяемых при колдовстве . Так, в « Метаморфозах » Апулея содержится рассказ о том, как колдунья убила бросившего её мужчину и при этом использовала губку в качестве средства, позволившего отсрочить момент наступления смерти (чтобы отвести подозрения от истинного убийцы) . Завёрнутая в шёлк губка с привязанной к ней нитью у древних евреев находила применение в качестве контрацептива .

Потребность в поставке губок многочисленным потребителям привела к развитию сбора губок как особого промысла , которым занимались . Они были столь сильными и атлетически сложенными, что ныряние за губками было включено в программу Олимпийских игр древности .
Древнюю традицию пронесли сквозь века жители островов Эгейского моря . В XIX и начале XX века Додеканесские острова (прежде всего остров Калимнос , в окрестностях которого находились большие популяции губок) превратились в крупнейший центр добычи морских губок и торговли ими . Трудный и нередко опасный промысел ныряльщиков стал важнейшей темой местного фольклора (в частности, многих песен ) . Первоначально ныряльщики Калимноса прыгали за борт лодки почти обнажёнными, держа в руках для быстрого погружения 15-килограммовый плоский камень; после того, как в 1865 году был изобретён водолазный скафандр , они стали использовать это новое снаряжение, позволившее увеличить глубину и длительность погружений. К концу XX века, однако, объёмы добычи и продажи морских губок значительно сократились, поскольку их вытесняют синтетические губки .
Помимо Греции , длительную историю имеет и добыча губок у побережья Сирии , Египта , Ливии и Туниса , где этот промысел также стал частью культурного наследия народов перечисленных стран. Коммерческое значение приобрёл сбор губок, принадлежащих к 5 видам родов Spongia и Hippospongia : S. officinalis , S. mollissima (его часто, впрочем, включают в состав вида S. officinalis ), , , . В 1841 году начались добыча морских губок с целью экспорта в районе Карибского моря — сначала на Багамах , несколько позже на Кубе и на побережье Флориды (район Ки-Уэст ); здесь к числу добываемых видов морских губок добавились , , , , и некоторые другие виды .

Один из экземпляров туалетной губки , получивший название , вошёл в число известных христианских реликвий . По свидетельству Евангелия от Иоанна , один из римских воинов, стерегущих распятого Христа , в ответ на его просьбу попить обмакнул губку в уксус и поднёс её к устам Иисуса, чтобы тот утолил жажду . Святая Губка, ставшая объектом почитания, хранилась сначала в Иерусалиме , потом — в Константинополе ; в настоящее время её фрагменты находятся в четырёх римских соборах ( Латеранская базилика , базилики Санта-Кроче-ин-Джерусалемме , Санта-Мария-Маджоре и Санта-Мария-ин-Трастевере ). Ещё один крупный фрагмент Святой Губки был вместе с несколькими другими реликвиями приобретён в 1239—1241 гг. французским королём Людовиком IX у латинского императора Балдуина II за огромную сумму и помещён в возведённую им парижскую капеллу Сент-Шапель . Небольшой фрагмент Святой Губки находится также в ковчеге Дионисия , хранящемся в Московском Кремле .

Изображение Святой Губки нередко появляется в произведениях изобразительного искусства . На фреске « Страшный суд » работы Микеланджело (1537—1541 гг.), находящейся на алтарной стене Сикстинской капеллы в Ватикане , можно видеть изображение ангела, держащего в руках шест со Святой Губкой на конце . Аналогичная фигура ангела со Святой Губкой встречается на фресках работы Франческо Приматиччо (1541—1544 гг.), украшающих своды хоров часовни Святой Марии в Шали (коммуна в департаменте Уаза на севере Франции ) . Скульптура ангела со Святой Губкой в руках стала наиболее известным произведением итальянского скульптора , ученика Лоренцо Бернини ; эта скульптора, изваянная Джорджетти в 1668 или 1669 году по заданию своего учителя, является одной из десяти скульптур ангелов, установленных на мосту Святого Ангела в Риме (выполненная Джорджетти в качестве эскиза терракотовая голова будущей скульптуры хранится в Эрмитаже ) .

Святая Губка (вместе со снятым с головы распятого Христа Терновым венцом ) фигурирует и в правом нижнем углу картины Питера Пауля Рубенса « Снятие с креста » (1612), которая представляет собой один из алтарных образов в кафедральном Соборе Антверпенской Богоматери .
Скелеты стеклянных губок , очищенные от органического вещества, используют как украшения и сувениры. Особенно ценится в этом плане губка Корзинка Венеры ( Euplectella ), скелет которой имеет форму нежного ажурного цилиндра с таким замысловатым строением, что кажется вырезанным рукою искусного мастера (в викторианскую эпоху один экземпляр данной губки был приобретён за весьма значительную по тем временам сумму в 5 гиней — свыше 500 фунтов в переводе на современные деньги) . В скелете этой губки выявлено семь иерархических структурных уровней, упорядоченная сборка которых позволяет преодолеть естественную хрупкость стекла и придать скелету значительную механическую прочность .
В Японии издавна существует обычай дарить молодожёнам на свадьбу экземпляр губки с находящейся внутри парой симбионтов — креветок вида (самцом и самкой, которые попадают внутрь губки личинками, а потом живут в ней всю свою жизнь: выбраться через мелкие поры губки они уже не могут) . Такой подарок символизирует нерушимую любовь и супружескую верность в соответствии с высказыванием «Вместе жить, вместе быть похороненными» .
В 1999 году американский телеканал « Nickelodeon » выпустил в показ мультсериал « Губка Боб Квадратные Штаны » ( англ. SpongeBob SquarePants ) режиссёра С. Хилленберга ; в сериале действующими лицами выступают представители морской фауны, а главным героем является губка Боб по фамилии «Квадратные Штаны» . В 2004 и 2015 годах телеканал выпустил полнометражные анимационные фильмы (соответственно, « Губка Боб Квадратные Штаны » и « Губка Боб Квадратные Штаны: Губка из воды »), ставшие продолжением мультсериала .
|
Классификация
животных
|
|
|---|---|
| Пластинчатые | |
| Гребневики | |
| Стрекающие | |
| Bilateria | |
|
|
Эта статья победила на конкурсе
статьи года
и была признана статьёй 2017 года русской Википедии.
|