Ядро Linux
- 1 year ago
- 0
- 0


Кле́точное ядро́ ( лат. nucleus ) — окружённая двумя мембранами важная структура ( органелла , компартмент ) эукариотической клетки . В клетках прокариот ядра нет. В клетках эукариот обычно одно ядро, однако некоторые типы клеток, например, эритроциты млекопитающих , не имеют ядра, а другие содержат несколько ядер.
В ядре заключена бо́льшая часть генетического материала клетки, представленного хромосомами , длинными линейными молекулами ДНК , связанными с белками . Генетический материал, локализованный в хромосомах, составляет ядерный геном . Ядро поддерживает целостность генетического материала, а входящие в его состав структуры управляют клеточными процессами, регулируя экспрессию генов , поэтому ядро является, по сути, контролирующим центром клетки. К основным структурам, из которых состоит ядро, относят хроматин , ядрышко , ядерную оболочку — двойную мембрану, окружающую ядро и изолирующую его от цитоплазмы , а также ядерный матрикс , который включает ядерную ламину — сеть филаментов , обеспечивающая механическую поддержку ядра, подобно цитоскелету в цитоплазме.
Поскольку ядерная оболочка непроницаема для крупных молекул, транспорт молекул через ядерную оболочку ( ) обеспечивают ядерные поры . Поры пронизывают обе ядерные мембраны и формируют сквозной канал, через который малые молекулы и ионы проходят свободно, а крупные молекулы активно транспортируются с участием белков-переносчиков. Перенос через ядерные поры таких крупных молекул, как белки и РНК , необходим для экспрессии генов, поддержания хромосом и сборки рибосомных субъединиц. Хотя внутри ядра нет окружённых мембраной субкомпартментов, его внутреннее содержимое неоднородно и содержит ряд ядерных телец, которые состоят из особых белков, молекул РНК и частей хромосом. Самое известное ядерное тельце — ядрышко , в котором происходит сборка рибосомных субъединиц. После образования в ядрышке рибосомные субъединицы транспортируются в цитоплазму, где они осуществляют трансляцию мРНК .

Ядро стало первой из органелл , открытых учёными-естествоиспытателями в составе клетки. Самые ранние рисунки клеток и их ядер принадлежат основоположнику научной микроскопии Антони ван Левенгуку (1633—1723), который наблюдал ядро в эритроцитах лосося . Описания ядра также выполнил в 1802 году , а более детальное описание было выполнено в 1831 году шотландским ботаником Робертом Броуном и представлено на собрании Лондонского Линнеевского общества . Броун изучал орхидеи под микроскопом и обнаружил в клетках наружного слоя цветка непрозрачные области, которые он называл «ареолами» или «ядрами» .
Броун не делал предположений относительно функций ядра. В 1838 году Маттиас Шлейден предположил, что ядро участвует в образовании новых клеток, поэтому он ввёл для обозначения ядер термин «цитобласт» (клеточный строитель). Он был уверен, что наблюдал сборку новых клеток вокруг «цитобластов». Убеждённым оппонентом этого взгляда был Франц Мейен , описавший клетки, размножающиеся посредством деления , и считавший, что у многих клеток может не быть ядра. Идея об образовании клеток , то есть с нуля, посредством цитобластов или иначе, противоречила работам Роберта Ремака (1852) и Рудольфа Вирхова (1855), которые окончательно утвердили новую парадигму, утверждающую, что клетки могут образовываться только из клеток («Omnis cellula e cellula»). Функции ядра оставались неясными .
Между 1877 и 1878 годами Оскар Гертвиг опубликовал несколько работ по оплодотворению яиц у морских ежей , в которых показал, что при оплодотворении ядро сперматозоида проникает внутрь яйцеклетки и сливается с её ядром. Впервые было показано, что новая особь развивается из единственной клетки, имеющей ядро. Это противоречило теории Эрнста Геккеля , согласно которой в ходе эмбрионального развития особи последовательно проходятся все этапы филогении её вида , а потому, в частности, поколение первых клеток с ядром якобы образуется из «монерулы» — бесструктурной массы первичной слизи. В связи с этим необходимость ядра сперматозоида для оплодотворения некоторое время была предметом дискуссий. Однако Гертвиг подтвердил свои наблюдения исследованиями на других животных, включая земноводных и моллюсков . В 1884 году Эдуард Страсбургер показал то же самое для растений. Это проложило путь к гипотезе о том, что ядро передаёт наследственный материал. В 1873 году Август Вейсман высказал идею о равнозначности материнского и отцовского материала для наследственности. Функция ядра как носителя генетической информации стала очевидной лишь позже, после открытия митоза и открытия заново законов Менделя в начале XX столетия. На основании этих открытий была сформулирована хромосомная теория наследственности .
Ядро — крупнейшая органелла животных клеток . У млекопитающих диаметр ядра составляет примерно 6 мкм , а само ядро составляет около 10 % объёма клетки . Вязкая жидкость, заполняющая ядро, называется нуклеоплазмой и по химическому составу близка к цитозолю , окружающему ядро .

|

|
Ядерная оболочка состоит из двух мембран (наружной и внутренней), которые расположены параллельно на расстоянии от 10 до 50 нм . Ядерная оболочка полностью окружает ядро, отделяя генетический материал клетки от цитоплазмы и служа барьером, предотвращающим свободную диффузию макромолекул между нуклеоплазмой и цитоплазмой . Наружная ядерная мембрана продолжается в мембрану шероховатого эндоплазматического ретикулума (ЭПР) и покрыта рибосомами . Промежуток между ядерными мембранами называется перинуклеарным пространством и продолжается в люмен ЭПР .
Ядерные поры, представляющие собой заполненные водой каналы в ядерной оболочке , состоят из множества белков, называемых нуклеопоринами . У человека масса пор составляет около 120 000 кДа ,что в 40 раз больше массы рибосомы ; при этом у дрожжей в состав ядерных пор входит около 50 белков, а у позвоночных — несколько сотен . Хотя диаметр пор составляет 100 нм , ширина щели, через которую могут проходить молекулы, из-за наличия внутри пор регуляторных систем составляет всего 9 нм . В такую щель могут проходить водорастворимые малые молекулы, но не крупные молекулы — такие, как нуклеиновые кислоты и большие белки; для переноса этих молекул в ядро необходим активный (то есть энергозатратный) транспорт. На оболочке ядра типичной клетки млекопитающего располагается от 3000 до 4000 пор , и у каждой в месте слияния двух ядерных мембран находится кольцевая структура, имеющая 8 осей симметрии . К кольцу прикрепляется особая структура, известная как ядерная корзина, которая выдаётся в нуклеоплазму, а несколько её филаментов выдаются в цитоплазму. Обе структуры необходимы для опосредования связывания транспортных ядерных белков .
Большинство белков, субъединицы рибосом и некоторые ДНК переносятся через ядерные поры посредством семейства транспортных факторов, известных как . Кариоферины, опосредующие транспорт в ядро, также называются , а опосредующие транспорт из ядра — экспортинами. Большинство кариоферинов непосредственно взаимодействуют со своим грузом, но некоторые используют для этого белки . Стероидные гормоны (такие, как кортизол и альдостерон ), а также другие жирорастворимые малые молекулы могут диффундировать в цитоплазму внутрь клетки через клеточную мембрану; в цитоплазме они связываются с белковыми ядерными рецепторами, которые доставляют их в ядро. Здесь ядерные рецепторы , связанные со своими лигандами, функционируют как транскрипционные факторы, а в отсутствие лиганда многие рецепторы функционируют как гистондеацетилазы , подавляющие экспрессию некоторых генов .

В клетках животных механическую поддержку ядра обеспечивают две сети из промежуточных филаментов : ядерная ламина, представляющая собой сеть промежуточных филаментов на внутренней поверхности ядра, а также менее организованные филаменты на цитозольной поверхности ядра. Обе системы филаментов обеспечивают поддержку ядра и служат для закрепления хромосом и ядерных пор .
Ядерная ламина состоит в основном из белков, известных как ламины . Как и все белки, ламины синтезируются в цитоплазме и далее транспортируются внутрь ядра, где они вставляются в ядерную ламину . Расположенные на наружной стороне ядерной оболочки белки (такие, как ), связываются с элементами цитоскелета, что обеспечивает структурную поддержку ядру. Ламины также обнаруживаются в нуклеоплазме, где они образуют другую регулярную структуру, известную как нуклеоплазматическая вуаль ( англ. nucleoplasmic veil ) ; последнюю можно визуализировать с использованием флуоресцентной микроскопии . Функция вуали неизвестна, но известно, что её нет в ядрышке и она присутствует в интерфазе клеточного цикла . Входящие в состав вуали ламины (такие, как LEM3) связываются с хроматином , и нарушения в их структуре подавляют транскрипцию белоккодирующих генов .
Как и другие белки промежуточных филаментов, мономеры ламинов содержат α-спиральный домен , используемый двумя мономерами, чтобы обвиться вокруг друг друга, образуя димер , имеющий структуру . Два димера далее связываются своими боковыми сторонами в антипараллельной ориентации, образуя тетрамер, известный как протофиламент. Восемь тетрамеров объединяются в скрученный, похожий на верёвку филамент. Филаменты могут собираться и разбираться динамическим образом, то есть длина филамента зависит от относительных скоростей его сборки и разборки .

В ядре находится большая часть генетического материала клетки, представленного линейными молекулами ДНК, которые организованы в структуры, известные как хромосомы . Суммарная длина молекул ДНК клетки человека составляет около 2 м . В течение интерфазы клеточного цикла данные молекулы в комплексе с белками формируют так называемый ядерный хроматин , а при клеточном делении хромосомы конденсируются и предстают в виде отдельных микроскопически различимых образований. Небольшое количество внеядерного клеточного генетического материала располагается в митохондриях и, в случае растительной клетки , в хлоропластах .
Известно два вида хроматина. В эухроматине ДНК наименее плотно организована; он содержит гены, которые транскрибируются наиболее часто . Другой вид хроматина, гетерохроматин , более компактен и содержит ДНК, транскрибируемую редко или никогда. Гетерохроматин подразделяется на факультативный, который формируется в части клеток в процессе развития , и конститутивный, присутствующий во всех клетках на всех стадиях развития и находящийся в основном в теломерных и околоцентромерных участках хромосом . В течение интерфазы хроматин каждой хромосомы занимает свою область ядра — хромосомную территорию , то есть, хроматин разных хромосом не перемешивается . Активные гены, которые, как правило, располагаются в эухроматине, обычно располагаются на границе хромосомной территории .
В ядре клеток млекопитающих содержится ряд дискретных субкомпартментов , которые называются ядерными тельцами. Они осуществляют компартментализацию ядра, создавая внутри него отдельные пространства, которые обладают определёнными свойствами. Многие ядерные тельца осуществляют специфические функции — например, синтез и процессинг пре-рибосомных РНК в ядрышке, накопление и сборку компонентов сплайсосом в спеклах (см. ниже) или накопление молекул РНК в параспеклах . Механизмы, которые обеспечивают выполнение ядрышковыми тельцами этих функций, очень разнообразны. В некоторых случаях ядерное тельце может служить местом протекания определённых процессов, например, транскрипции. В других случаях ядерные тельца, по-видимому, опосредованно регулируют локальные концентрации своих компонентов в нуклеоплазме. Подобно цитоплазматическим органеллам, ядерные тельца содержат специфический набор белков, которые определяют их структуру на молекулярном уровне. Однако, в отличие от органелл цитоплазмы, ядерные тельца не окружены липидными мембранами, и их структурная целостность целиком обеспечивается белок-белковыми и РНК-белковыми взаимодействиями. Ниже в таблице перечислены основные характеристики ядерных телец .
| Ядерное тельце | Функции | Характерные компоненты | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|---|
| Ядрышко | Биогенез рибосом | Машинерия , факторы процессинга рРНК и сборки рибосомных субъединиц | 3—8 | 1—4 |
| Спеклы | Накопление и сборка факторов сплайсинга | Факторы сплайсинга пре-мРНК | 2—3 | 20—50 |
| Стрессовые ядерные тельца | Регуляция транскрипции и сплайсинга в условиях стресса | , HAP | 1—2 | 3—6 |
| Тельце гистоновых локусов | Процессинг пре-мРНК гистонов | , FLASH, мяРНП | 0,2—1,2 | 2—4 |
| Тельце Кахаля | Биогенез, созревание и кругооборот малых РНК | Коилин , | 0,2—1,5 | 1—10 |
| PML-тельце | Регуляция стабильности генома, репарация ДНК , контроль транскрипции, защита от вирусов | PML | 0,1—1 | 10—30 |
| Параспеклы | Регуляция мРНК, редактирование РНК | Некодирующие РНК NEAT1/MENε/β, белки PSP1, p54 nrb /NONO | 0,2—1 | 2—20 |
| Околоядрышковый компартмент | Посттранскрипционная регуляция набора РНК, синтезированных РНК-полимеразой III | PTB | 0,2—1 | 1—2 |

Ядрышко — это отдельная плотная структура в ядре. Она не окружена мембраной и формируется в области расположения рДНК — тандемных повторов генов рибосомной РНК (рРНК) , называемых ядрышковыми организаторами . Главная функция ядрышка — синтез рРНК и образование рибосом. Структурная целостность ядрышка зависит от его активности, и инактивация генов рРНК приводит к смешению ядрышковых структур .
На первой стадии образования рибосом фермент РНК-полимераза I транскрибирует рДНК и образует пре-рРНК, которая далее разрезается на 5,8S, 18S и 28S рРНК . Транскрипция и посттранскрипционный процессинг рРНК происходят в ядрышке при участии малых ядрышковых РНК (snoРНК), некоторые из которых происходят из сплайсированных интронов мРНК генов, кодирующих белки, связанные с работой рибосом. Собранные рибосомные субъединицы — это самые крупные структуры, проходящие через ядерные поры .
При рассматривании под электронным микроскопом в ядрышке можно выделить три компонента: фибриллярные центры (ФЦ), окружающий их плотный фибриллярный компонент (ПФК) и гранулярный компонент (ГК), который, в свою очередь, окружает ПФК. Транскрипция рРНК происходит в ФЦ и на границе ФЦ и ПФК, поэтому при активации образования рибосом ФЦ становятся хорошо различимы. Разрезание и модификации рРНК происходят в ПФК, а последующие этапы образования рибосомных субъединиц, включающие загрузку рибосомных белков, происходят в ГК .

Тельце Кахаля (ТК) — ядерное тельце, имеющееся у всех эукариот. Оно идентифицируется по наличию сигнатурного белка коилина и специфических РНК (scaРНК). В ТК также содержится белок SMN ( англ. survival of motor neurons ). В ТК наблюдается высокая концентрация сплайсирующих малых ядерных рибонуклеопротеинов (мяРНП) и других факторов процессинга РНК, поэтому считается, что ТК служат местами сборки и/или посттранскрипционной модификации факторов сплайсинга. ТК присутствует в ядре во время интерфазы, но исчезает в митозе. В биогенезе ТК прослеживаются свойства самоорганизующейся структуры .
Когда внутриклеточная локализация SMN впервые изучалась методом иммунофлуоресценции , то белок обнаруживался во всей цитоплазме, а также в ядрышковом тельце, сходном по размеру с ТК и часто расположенном рядом с ТК. По этой причине данное тельце было названо «близнецом ТК» ( англ. gemini of CB ) или просто gem. Однако оказалось, что линия клеток HeLa , в которой было открыто новое тельце, была необычной: в других линиях клеток человека, а также у плодовой мушки Drosophila melanogaster SMN колокализовался с коилином в ТК. Поэтому в общем случае SMN можно рассматривать как важный компонент ТК, а не как маркер отдельного ядерного тельца .
Тельце гистоновых локусов ( англ. histone locus body, HLB ) содержит факторы, необходимые для процессинга пре-мРНК гистонов. Как и следует из названия, тельца гистоновых локусов ассоциированы с генами, кодирующими гистоны; поэтому предполагается, что в тельцах гистоновых локусов концентрируются факторы сплайсинга. Тельце гистоновых локусов присутствует в клетке во время интерфазы и исчезает с наступлением митоза. Тельце гистоновых локусов нередко рассматривается вместе с тельцем Кахаля по нескольким причинам. Во-первых, в некоторых тельцах гистоновых локусов содержится маркер телец Кахаля — коилин. Во-вторых, эти тельца нередко физически находятся рядом, поэтому между ними наблюдается некоторое взаимодействие. Наконец, очень крупные тельца Кахаля ооцитов земноводных обладают свойствами обоих телец .
Тельца промиелоцитной лейкемии ( англ. Promyelocytic leukaemia bodies ), или PML-тельца — сферические тельца, разбросанные по всей нуклеоплазме и достигающие около 0,1—1,0 мкм в диаметре. Они известны также под такими названиями, как ядерный домен 10 ( англ. nuclear domain 10 (ND10) ), тельца Кремера ( англ. Kremer bodies ) и онкогенные домены PML ( англ. PML oncogenic domains ). Тельца PML названы по одному из своих ключевых компонентов — белку промиелоцитной лейкемии (PML). Они часто наблюдаются ассоциированными с тельцами Кахаля и тельцами деления ( англ. cleavage body ) . PML-тельца принадлежат ядерному матриксу и могут быть задействованы в таких процессах, как репликация ДНК , транскрипция и эпигенетический сайленсинг генов . Ключевым фактором организации этих телец выступает белок PML, который привлекает другие белки; последние, по современным представлениям, объединены лишь тем, что они SUMOилированы . Мыши , у которых ген PML делетирован , лишены PML-телец, однако развиваются и живут нормально, поэтому PML-тельца не выполняют незаменимых биологических функций .
Спеклы ( англ. speckle ) — это ядерные тельца, которые содержат факторы сплайсинга пре-мРНК и располагаются в интерхроматиновых участках нуклеоплазмы клеток млекопитающих. При флуоресцентной микроскопии спеклы выглядят как пятнистые тельца неправильной формы, различных размеров, а при электронной микроскопии они выглядят как кластеры интерхроматиновых гранул. Спеклы — динамические структуры, и содержащиеся в них белки и РНК могут перемещаться между спеклами и другими ядерными тельцами, включая участки активной транскрипции. На основании исследований состава, структуры и поведения спеклов была создана модель, объясняющая функциональную компартментализацию ядра и организацию машинерии экспрессии генов , сплайсирующих малые ядерные рибонуклеопротеины и другие белки, необходимые для сплайсинга пре-мРНК . Из-за изменяющихся потребностей клетки состав и расположение спеклов изменяется согласно транскрипции мРНК и посредством регуляции фосфорилирования специфических белков . Сплайсирующие спеклы также известны как ядерные спеклы, компартменты сплайсирующих факторов, кластеры интерхроматиновых гранул и B-снурпосомы ( англ. B snurposomes ) . B-снурпосомы найдены в ядрах ооцитов земноводных и зародышах плодовой мушки Drosophila melanogaster . На электронных микрофотографиях B-снурпосомы предстают прикреплёнными к тельцам Кахаля или отдельно от них. Кластеры интерхроматиновых гранул служат местами скопления факторов сплайсинга .

Параспеклы — это ядерные тельца неправильной формы, располагающиеся в интерхроматиновом пространстве ядра . Впервые они были описаны у клеток HeLa, у которых имеется 10—30 параспеклов на ядро, но сейчас параспеклы обнаружены во всех первичных клетках человека, в клетках трансформированных линий и на срезах тканей . Своё название они получили из-за своего расположения в ядре — вблизи спеклов .
Параспеклы — динамические структуры, которые изменяются в ответ на изменения в метаболической активности клетки. Они зависят от транскрипции , и в отсутствие транскрипции, проводимой РНК-полимеразой II , параспеклы исчезают, а все входящие в их состав белки (PSP1, p54nrb, PSP2, CFI(m)68 и PSF) формируют серповидный околоядрышковый кэп . Этот феномен наблюдается в ходе клеточного цикла: параспеклы присутствуют в интерфазе и всех фазах митоза, за исключением телофазы . В ходе телофазы формируются дочерние ядра, и РНК-полимераза II ничего не транскрибирует, поэтому белки параспеклов и формируют околоядрышковый кэп . Параспеклы участвуют в регуляции экспрессии генов, накапливая те РНК, где есть двухцепочечные участки, которые подвергаются редактированию, а именно превращению аденозина в инозин . Благодаря этому механизму параспеклы задействованы в контроле экспрессии генов при дифференцировке , вирусной инфекции и стрессе .
Околоядрышковый компартмент (ОК) — ядерное тельце неправильной формы, которое характеризуется тем, что располагается на периферии ядрышка. Несмотря на физическую связь, эти два компартмента структурно различны. Обычно ОК обнаруживают в клетках злокачественных опухолей . ОК — динамическая структура, и содержит очень много РНК-связывающих белков и РНК-полимеразу III. Структурная стабильность ОК обеспечивается транскрипцией, осуществляемой РНК-полимеразой III, и наличием ключевых белков. Поскольку присутствие ОК обычно связано со злокачественностью и со способностью к метастазированию , их рассматривают как потенциальные маркеры рака и других злокачественных опухолей. Показана ассоциация ОК со специфическими локусами ДНК .
Стрессовые ядерные тельца формируются в ядре при тепловом шоке. Они образуются при непосредственном взаимодействии транскрипционного фактора теплового шока 1 ( ) и перицентрических тандемных повторов в последовательности сателлита III, что соответствует сайтам активной транскрипции некодирующих транскриптов сателлита III. Распространено мнение, что такие тельца соответствуют очень плотно упакованным формам рибонуклеопротеиновых комплексов. Считается, что в клетках, подвергающихся стрессу, они участвуют в быстрых, временных и глобальных изменениях в экспрессии генов посредством различных механизмов — например, ремоделирования хроматина и захватывания факторов транскрипции и сплайсинга. В клетках, находящихся в нормальных (не стрессовых) условиях, стрессовые ядерные тельца обнаруживаются редко, однако их количество резко увеличивается под действием теплового шока. Стрессовые ядерные тельца найдены только в клетках человека и других приматов .
Ядерные тельца-сироты ( англ. orphan nuclear bodies ) — нехроматиновые ядерные компартменты, которые исследованы гораздо хуже, чем другие хорошо охарактеризованные структуры ядра. Некоторые из них выступают как места, в которых белки модифицируются белками SUMO и/или происходит протеасомная деградация белков, помеченных убиквитином . Ниже в таблице приведены характеристики известных ядерных телец-сирот .
| Ядерное тельце | Описание | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|
| Кластосома | Концентрирует протеасомные комплексы 20S и 19S и белки, связанные с убиквитином. Обнаруживается, главным образом, тогда, когда стимулируется активность протеасом, и разбирается при ингибировании активности протеасом. | 0,2—1,2 | 0—3 |
| Тельце деления ( англ. cleavage body ) | Обогащено факторами деления и , а также белком , содержащим . Обнаруживается в основном в S-фазе , ингибирование транскрипции на него не влияет. | 0,2—1,0 | 1—4 |
| Домен OPT | Обогащён факторами транскрипции и PTF. Частично колокализуется с сайтами транскрипции. Обнаруживается в основном в поздней G1-фазе , разбирается при ингибировании транскрипции. | 1,0—1,5 | 1—3 |
| Тельце Polycomb | Обнаруживается в клетках человека и дрозофилы, обогащено белком PcG . У человека накапливает белки RING1 , , HPC, может быть связано с околоцентромерным гетерохроматином. | 0,3—1,0 | 12—16 |
| Тельце Sam68 | Накапливает белок Sam68 и схожие с ним белки SLM-1 и SLM-2. Разбирается при ингибировании транскрипции. Вероятно, обогащено РНК. | 0,6—1,0 | 2—5 |
| Тельце SUMO | Обогащено белками SUMO и SUMO-конъюгирующим ферментом . Концентрирует транскрипционные факторы p CREB , CBP , . | 1—3 | 1—3 |
Ядерная оболочка защищает ДНК клетки и участвует в гораздо более сложной регуляции экспрессии генов по сравнению с прокариотической клеткой. У прокариот транскрипция и трансляция являются сопряжёнными процессами и трансляция мРНК в белок начинается ещё до того, как она будет полностью синтезирована. В клетках эукариот цитоплазма, в которой проходит трансляция, и транскрипция, протекающая в ядре, пространственно разобщены, поэтому возникает необходимость в обеспечении транспорта молекул между ядром и цитоплазмой .
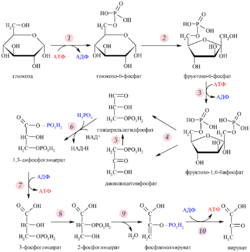
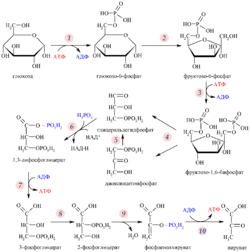
Ядерная оболочка даёт ядру возможность контролировать своё содержимое и отделяет его от остальной цитоплазмы. Это имеет важное значение для регуляции процессов, протекающих по обе стороны ядерной оболочки. Когда цитоплазматический процесс должен быть как-то ограничен, то обычно его ключевой участник переносится в ядро, где он взаимодействует с факторами транскрипции и таким образом запускает подавление образования некоторых ферментов, задействованных в цитоплазматическом процессе. Например, такой регуляторный механизм имеется у гликолиза — процесса, в ходе которого клетка извлекает энергию из молекулы глюкозы . Первую реакцию гликолиза осуществляет фермент гексокиназа , преобразуя молекулу глюкозы в глюкозо-6-фосфат . Когда концентрация фруктозо-6-фосфата (вещества, в ходе гликолиза образующегося из глюкозо-6-фосфата) возрастает, регуляторный белок отправляет гексокиназу в ядро , где она формирует транскрипционный репрессирующий комплекс, который подавляет экспрессию генов, кодирующих ферменты гликолиза .
Чтобы контролировать, какие именно гены транскрибируются, в клетке транскрипционные факторы не имеют физического доступа к ДНК, пока они не будут активированы в ходе определённого сигнального пути . Это предотвращает даже низкую экспрессию неправильных генов. В частности, в случае контролируемых NF-κB генов, которые принимают участие в воспалительном процессе , транскрипция индуцируется под действием сигнального пути, например, начинающегося со связывания сигнальной молекулы TNF-α со своим рецептором на клеточной мембране и в конце концов приводящего к активации фактора транскрипции NF-κB. Сигнал ядерной локализации , имеющийся у NF-κB, позволяет ему проходить в ядро и из него через ядерные поры; в ядре он стимулирует транскрипцию генов-мишеней .
Компартментализация предотвращает транскрипцию клеткой несплайсированной мРНК. Эукариотические мРНК содержат интроны, которые должны быть удалены до того, как начнётся трансляция мРНК. Сплайсинг, то есть удаление интронов, протекает в ядре, что предотвращает доступ к пре-мРНК рибосом, находящихся вне ядра. Если бы ядра не было, то рибосомы начинали бы транслировать незрелые мРНК, что привело бы к образованию неправильных белковых продуктов .
Поскольку транскрипция протекает в ядре, ядро содержит множество белков, непосредственно участвующих в транскрипции или регулирующих этот процесс. К этим белкам относятся хеликазы , которые расплетают двойную спираль ДНК, облегчая доступ к ней других белков, РНК-полимеразы , которые синтезируют РНК, топоизомеразы , влияющие на топологию ДНК, а также разнообразные факторы транскрипции .

Выход из ядра и вход в ядро крупных молекул контролируется ядерными порами. Хотя малые молекулы могут проникать в ядро без всякой регуляции, макромолекулы — такие, как белки и РНК — должны связаться с кариоферинами для транспорта в ядро (импортинами) и из ядра (экспортинами). Белки, которые должны быть транспортированы из цитоплазмы в ядро, содержат особую аминокислотную последовательность, известную как сигнал ядерной локализации, с которой связываются импортины. Аналогичным образом белки, которые должны выйти из ядра, содержат , распознаваемый экспортинами. Способность импортинов и экспортинов переносить свой груз регулируется ГТФазами — ферментами, которые гидролизуют ГТФ с высвобождением энергии . Ключевая ГТФаза ядерного транспорта — , которая может связываться с ГТФ или ГДФ , в зависимости от своего местонахождения (в ядре или в цитоплазме). В ядре взаимодействие Ran-ГТФ с импортином вызывает конформационные изменения в последнем, так что он отделяется от переносимого груза. Образованный комплекс Ran-ГТФ и импортина транспортируется в цитоплазму, где белок RanBP отделяет Ran-ГТФ от импортина. Отделение от импортина позволяет белку связаться с Ran-ГТФ и катализировать гидролиз ГТФ до ГДФ. Далее комплекс Ran-GDP распознаётся белком , который возвращает его в нуклеоплазму. В ядре белок заменяет ГДФ на ГТФ, образуя Ran-ГТФ и замыкая цикл .
Ядерный экспорт осуществляется похожим образом. В ядре экспортин связывается с белком-грузом и Ran-ГТФ и переносится через ядерную пору в цитоплазму, где комплекс диссоциирует . Ran-ГТФ гидролизует ГТФ до ГДФ под действием GAP, и комплекс Ran-ГДФ переносится в ядро, где ГДФ заменяется на ГТФ . Для транспорта через ядерную оболочку зрелых мРНК и тРНК также существуют специальные белки .
В течение жизни клетки ядро может быть разобрано (при делении клетки или при апоптозе ). В ходе этих процессов структурные компоненты ядра — ядерная оболочка и ядерная ламина — разрушаются. В большинстве клеток разборка ядра наблюдается в профазе митоза. Однако разборка ядра не приурочена строго к митозу и происходит не во всех клетках. Некоторые одноклеточные эукариоты (например, дрожжи ) подвергаются так называемому закрытому митозу, при котором ядерная оболочка остаётся целой. При закрытом митозе хромосомы перемещаются к разным сторонам ядра, которое потом делится надвое. Клетки высших эукариот, напротив, обычно подвергаются открытому митозу, в ходе которого ядерная оболочка распадается. Хромосомы мигрируют к разным полюсам веретена деления , и вокруг них заново формируются два ядра. Ядерная ламина тоже подвергается разборке из-за фосфорилирования ламинов такими киназами , как . Сборка ядерной ламины в дочерних ядрах начинается после дефосфорилирования ламинов .
Апоптоз — это контролируемый процесс разрушения клеточных компонентов, приводящего к гибели клетки. Перемены, связанные с апоптозом, происходят непосредственно с ядром и его содержимым. К их числу относится конденсация хроматина, а также дезинтеграция ядерной оболочки и ядерной ламины. Разрушение сети ламинов происходит с участием апоптотических протеаз , известных как каспазы , которые разрушают ламины и, таким образом, влияют на структурную целостность ядра. Разрушение ламинов иногда используется в качестве индикатора активности каспаз в исследованиях, посвящённых апоптозу. Клетки, в которых экспрессируются мутантные ламины, устойчивые к действию каспаз, при апоптозе не утрачивают целостность ядра, поэтому ламины играют ключевую роль в начале изменений, которое претерпевает ядро при апоптозе . Кроме того, ингибирование сборки ламинов в сеть запускает апоптоз .
Размеры, формы и морфология ядер эукариот изменяются в самых широких пределах. Если у пироплазмид и лейшманий диаметр ядра составляет 1—3 мкм , то у некоторых радиолярий ядра в диаметре достигают 400 мкм и даже 1 мм . Как правило, форма ядра у большинства эукариот близка к сферической, но иногда она способна принимать довольно причудливые очертания (это, в частности, относится к макронуклеусам инфузорий). Хотя у всех эукариот оболочка ядра состоит из двух мембран, число пор в ней у различных видов сильно варьирует, причём иногда к ней (как снаружи, так и изнутри) могут примыкать дополнительные слои; например, у многих свободноживущих амёб к внутренней стороне оболочки прилегает фиброзный слой с ячеистым строением, который значительно превосходит ядерную оболочку по толщине, а у радиолярий с внешней стороны оболочки располагаются дополнительные фибриллярные слои .
Значительным своеобразием отличается организация ядра у протистов из типа динофлагеллят (Dinoflagellata). Большинство их представителей имеет ядро, в котором хромосомы на протяжении всего клеточного цикла (в том числе и в интерфазе ) и практически лишены гистонов . Такой тип ядра получил название динокарион . При этом количество ДНК в динокарионе в десятки и сотни раз превосходит количество ДНК, приходящееся на клетку у представителей других групп эукариот . Впрочем, некоторые динофлагелляты ( Noctiluca , ) имеют обычные эукариотические ядра ; у других представителей типа в вегетативных клетках ядра обычные, а динокарион присутствует на других стадиях клеточного цикла (например, в гаметах) .

Клетки протистов обладают по крайней мере одним ядром . В то же время в организмах Metazoa встречаются и безъядерные клетки, которые, не имея ядра, утратили способность делиться с образованием двух дочерних клеток. Наиболее известным примером безъядерных клеток являются эритроциты млекопитающих, в которых отсутствуют и другие органеллы — такие, как митохондрии . Эритроциты созревают в костном мозге в процессе эритропоэза , в ходе которого они утрачивают ядра, другие органеллы и рибосомы. Ядро выталкивается из клетки при процессе дифференцировки эритробласта в ретикулоцит , который выступает непосредственным предшественником эритроцита . Под действием некоторых мутагенов в кровь могут выпускаться незрелые эритроциты, содержащие микроядра .
Большинство протистов имеет только одно ядро; у протистов, для которых характерен сложный жизненный цикл (например, у представителей типа апикомплексы (Apicomplexa) встречаются одноядерные и многоядерные стадии .
В ряде групп протистов клетки имеют несколько ядер на протяжении всей жизни; при этом многоядерные формы протистов способны достигать крупных размеров — порядка нескольких сантиметров в диаметре (в исключительных случаях — до метра и более) . Так, большинство представителей отряда дипломонад и, в частности, лямблии — хорошо известные паразиты кишечника млекопитающих и птиц из рода Giardia — имеют два функционально эквивалентных ядра, которые наследуются независимо в ходе митоза . У представителей рода (тип ) клетка содержит от 2 до 16 идентичных ядер. У жгутиконосцев из класса опалины (Opalinea) клетки также содержат несколько одинаковых ядер; их число существенно различается на различных стадиях жизненного цикла опалин. Много ядер у некоторых представителей отряда , причём количеству ядер соответствует и количество имеющихся в клетке .

В составе хлоропластов у криптофитовых и хлорарахниофитовых водорослей присутствует нуклеоморф — редуцированное ядро фототрофного эндосимбионта , инкорпорированного предками данных водорослей в ходе вторичного эндосимбиоза (у Cryptophyta происходила инкорпорация красной , а у Chlorarachnea — зелёной водоросли) .
У инфузорий и некоторых фораминифер наблюдается феномен ядерного дуализма, при котором в клетке присутствуют ядра двух типов: генеративный микронуклеус и вегетативный макронуклеус . При этом настоящий ядерный дуализм, при котором клетка содержит один или несколько мелких микронуклеусов и один или несколько крупных макронуклеусов, характерен для инфузорий и для определённых стадий (агамонтов) некоторых фораминифер (например, у Rotaliella heterokaryotica ) ; вообще же клетки или плазмодии фораминифер содержат от одного до нескольких тысяч ядер . В клетках инфузорий может быть как один, так и несколько микронуклеусов; это справедливо и для макронуклеусов. Микронуклеусы диплоидны , и именно в них происходит генетическая рекомбинация. Для макронуклеусов же характерен высокий уровень амплификации генов (так, у Paramecium tetraurelia уровень плоидности макронуклеуса составляет 1000—2000); впрочем, у инфузорий из класса микро- и макронуклеусы содержат почти одинаковый диплоидный набор ДНК. Макронуклеусы ответственны за клеточный метаболизм и являются местом синтеза РНК. В ходе деления клетки старые макронуклеусы обычно дегенерируют, новые же развиваются путём модификации микронуклеусов . Дифференцировка ядер на генеративные и вегетативные имеет место также у миксоспоридий (Myxosporea) и большинства акантарий (Acantharea); у последних такая дифференцировка происходит перед инцистированием : одно полиплоидное ядро даёт начало сначала вегетативным ядрам, а затем — генеративным, число которых в клетке в результате неоднократных делений достигает сотен .
Распространено и наличие двух ядер в клетках мицелия у грибов (особенно у образующих микоризу ) и в клетках сближаемых в современных классификациях с грибами микроспоридий . Данное явление известно как дикарион , или диплокарион . Встречающиеся у многих грибов несептированные гифы также, по существу, представляют собой гигантские многоядерные клетки .
У семенных растений тоже возможно появление многоядерных клеток. Например, многоядерную стадию развития проходят клетки эндосперма покрытосеменных (после двойного оплодотворения ) и женского гаметофита голосеменных (после мейоза ). В ряде случаев появление тканей с многоядерными клетками оказывается следствием механического или биохимического воздействия на организм растения-хозяина, оказанного паразитирующими на нём насекомыми . У многих покрытосеменных многоядерными являются клетки — слоя в пыльнике , ответственного за снабжение пыльцевых зёрен питательными веществами .
У человека и других позвоночных клетки скелетной мускулатуры ( миоциты ) сливаются с образованием многоядерного синцития . В нём ядра оттеснены к периферии, что даёт возможность занять внутреннее пространство сократимыми миофибриллами . Многоядерными также являются остеокласты — клетки костной ткани позвоночных, ответственные за её резорбцию ; в норме у млекопитающих они содержат от 2 до 30 ядер (в среднем — от 3 до 10), а при некоторых болезнях, которым сопутствует увеличение костной резорбции (при синдроме Педжета — Шрёттера , ревматоидном артрите и др.), остеокласты увеличиваются в размерах и число ядер в них возрастает (при синдроме Педжета — Шрёттера они могут содержать до 100 ядер) . Многоядерные клетки у человека и животных могут образовываться и при других патологических процессах. Так, слияние макрофага и моноцита с образованием гигантских многоядерных клеток происходит при воспалении , а также может говорить об образовании опухоли .
Клеточное ядро является важнейшей чертой эукариотических организмов, отличающей их от бактерий и архей . Несмотря на значительный прогресс в цитологии и молекулярной биологии, происхождение ядра не выяснено и является предметом научных споров. Выдвинуто 4 основных гипотезы происхождения клеточного ядра, но ни одна из них не получила широкой поддержки .
Гипотеза, известная как синтропная модель, предполагает, что ядро возникло в результате симбиотических взаимоотношений между археей и бактерией (ни археи, ни бактерии не имеют оформленных клеточных ядер). По этой гипотезе, симбиоз возник, когда древняя архея (сходная с современными метаногенными археями ), проникла в бактерию (сходную с современными миксобактериями ). Впоследствии архея редуцировалась до клеточного ядра современных эукариот. Эта гипотеза аналогична практически доказанным теориям происхождения митохондрий и хлоропластов , которые возникли в результате эндосимбиоза прото-эукариот и аэробных бактерий . Как свидетельство в пользу данной гипотезы рассматривается наличие у эукариот и архей одинаковых генов (в частности, генов гистонов ). Кроме того, миксобактерии быстро передвигаются, могут образовывать многоклеточные структуры и имеют киназы и G-белки , близкие к эукариотическим .

Согласно второй гипотезе, прото-эукариотическая клетка эволюционировала из бактерии без стадии эндосимбиоза. Доказательством модели является существование современных бактерий группы Planctomycetes , которые имеют ядерные структуры с примитивными порами и другие клеточные компартменты, ограниченные мембранами (ничего похожего у других прокариот не обнаружено) .
Согласно гипотезе вирусного эукариогенеза , окружённое мембраной ядро, как и другие эукариотические элементы, возникли вследствие инфекции прокариотической клетки вирусом. Это предположение основывается на наличии общих черт у эукариот и некоторых вирусов, а именно генома из линейных цепей ДНК, кэпирования мРНК и тесного связывания генома с белками ( гистоны эукариот принимаются аналогами вирусных ДНК-связывающих белков). По одной версии, ядро возникло при фагоцитировании (поглощении) клеткой большого ДНК-содержащего вируса . По другой версии, эукариоты произошли от древних архей, инфицированных поксвирусами . Эта гипотеза основана на сходстве ДНК-полимеразы современных поксвирусов и эукариот . Также предполагается, что нерешённый вопрос о происхождении пола и полового размножения может быть связан с вирусным эукариогенезом .
Четвёртая, самая новая гипотеза, названная экзомембранной гипотезой, утверждает, что ядро произошло от одиночной клетки, которая в процессе эволюции выработала вторую внешнюю клеточную мембрану; первичная клеточная мембрана после этого превратилась в ядерную мембрану, и в ней образовалась сложная система поровых структур ( ядерных пор ) для транспорта клеточных компонентов, синтезированных внутри ядра .

Мутации , затрагивающие белки различных компонентов ядра, нередко приводят к заболеваниям. Так, мутации, которые затрагивают ламины, приводящие к нарушениям в сборке филаментов ядерной ламины, лежат в основе группы редких наследственных заболеваний, известных как . Наиболее изучена группа ламинопатий, выступающих под общим названием прогерия . У больных прогерией наблюдается преждевременное старение, однако биохимические основы такого фенотипа неясны .
Наличие в крови антител к некоторым белкам хроматина, например, нуклеосомным комплексам, обусловливает аутоиммунные заболевания — такие, как системная красная волчанка . Данные антитела известны под названием , и их наличие также может быть связано с рассеянным склерозом как частью общего расстройства иммунной системы . Как и в случае прогерии, биохимическая подоплёка таких симптомов неясна .
Мутации в белках ядрышка часто приводят к различным раковым заболеваниям . Если в ядрышке проявляются дефекты образования рибосом, то наблюдаются заболевания, известные как . Нарушения в других ядерных тельцах тоже могут приводить к болезням. Так, присутствие в ядре маленьких палочек часто выявляется в случаях . Это заболевание обусловлено мутациями в гене актина , и сами палочки состоят из мутантного актина и других белков цитоскелета .
В норме ядерная оболочка служит барьером, который препятствует проникновению в ядро различных вирусов. Некоторым вирусам для репликации и/или сборки необходимы белки, находящиеся внутри ядра. Сборка и репликация ДНК-содержащих вирусов (это, например, герпесвирусы ) происходит внутри ядра, и вирионы покидают его, отпочковываясь от внутренней ядерной мембраны. Этот процесс сопровождается разборкой ядерной ламины с обращённой к ядру стороны внутренней ядерной мембраны .
|
|
|
|---|---|
| Словари и энциклопедии |
|
| В библиографических каталогах |
|
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
|
Ядерная мембрана
/
Ядерная ламина |
|

|
|||||||
|
Ядрышко
|
|||||||||
| Прочее |
|
||||||||
|
Эндомембранная система
|
|
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |