Земноводные
- 1 year ago
- 0
- 0

Земново́дные , или амфи́бии ( лат. Amphibia ) , — класс четвероногих позвоночных животных , включающий в себя (в числе прочих) тритонов , саламандр , лягушек и червяг и насчитывающий около 8 700 современных видов . Земноводные обитают практически повсеместно (за исключением Антарктиды , Северного полюса и некоторых островов ). В последние десятилетия по всему миру наблюдается резкое сокращение популяций амфибий, в основном из-за истребления людьми, разрушения среды обитания, опасных болезней: под серьёзной угрозой исчезновения находятся более 3045 видов .
Самые ранние амфибии в широком смысле произошли в девонский период — более 370 млн лет назад — от лопастепёрых рыб с лёгкими, что помогло им приспособиться к суше. Земноводные диверсифицировались и стали доминирующими наземными позвоночными в каменноугольный период , но позже были вытеснены рептилиями и синапсидами . Происхождение современных амфибий — очень спорная тема. Однако складывается общее мнение, что они, вероятно, произошли от темноспондилов , самой разнообразной группы доисторических земноводных в пермский период .
Самая маленькая амфибия (и позвоночное животное) в мире — лягушка из Папуа—Новой Гвинеи Paedophryne amauensis длиной всего 7,7—8 мм. Самая крупная из ныне живущих амфибий — китайская исполинская саламандра длиной 1,8 м и массой 70 кг; вымершие прионозух и мастодонзавр достигали в длину 6—9 метров и весили около двух тонн. Современные же представители класса, как правило, невелики .
Земноводные, наряду с пресмыкающимися, являются традиционным объектом изучения герпетологии , однако иногда выделяют и самостоятельную науку — батрахологию .
Группа земноводных занимает промежуточное положение между наземными и водными позвоночными животными: размножение и развитие у большинства видов происходит в водной среде, а взрослые особи обитают на суше, тем самым относясь к анамниям . Амфибии являются важной частью экосистемы и употребляются в пищу многими более крупными хищниками .
В индивидуальном развитии земноводные, как правило, проходят стадию живущей в воде личинки (у бесхвостых называемой головастиком ). Личинки амфибий сохраняют многие черты строения, присущие рыбам , такие как наличие жабр , боковой линии , двухкамерного сердца . В ходе метаморфоза , головастики претерпевают значительные изменения: у них появляются конечности, жабры заменяются лёгкими , происходит перестройка кровеносной и выделительной систем . Большинство земноводных откладывают икру в воду; в то же время, некоторые виды откладывают икру на суше, а другие не проходят стадию головастика, являясь живородящими . Взрослые особи большинства видов являются хищниками, чья диета состоит из мелких беспозвоночных (крупные виды могут питаться и небольшими позвоночными ), но существуют также и всеядные виды и даже несколько видов, питающихся фруктами . Взрослые земноводные дышат лёгкими, также они могут дышать и водопроницаемой кожей, а у некоторых мелких саламандр и лягушек нет лёгких, и дышат они только кожей. Внешне они похожи на пресмыкающихся, таких как ящерицы или змеи , но, наряду с млекопитающими и птицами , рептилии являются амниотами , в отличие от амфибий .
Главные признаки земноводных — слизистая, гладкая и водопроницаемая кожа, период метаморфоза (превращения из личинки во взрослое животное), подвижные веки, хладнокровность и необходимость воды для размножения. Амфибии — экзотермные (холоднокровные) позвоночные, у которых температура тела не поддерживается за счёт внутренних физиологических процессов. Их скорость метаболизма низкая. У многих земноводных мускулистые языки; черепа современных амфибий в основном широкие и короткие, часто не полностью окостеневшие и содержащие большое количество хряща . Кожа земноводных слабо ороговевает и не покрыта чешуёй, за исключением некоторых червяг. Она содержит много слизистых желёз, а у некоторых видов ядовитые железы (разновидность зернистых желёз). Сердце амфибий трёхкамерное, состоит из двух предсердий и одного желудочка . Амфибии, ведущие пресноводный образ жизни, в качестве продуктов азотистого обмена выделяют аммиак (аммониотеличный обмен), а наземные формы — мочевину (уреотеличный обмен). Большинство земноводных откладывают яйца в воду .
Слово «амфибия» происходит от франц. amphibie, далее от др.-греч. ἀμφίβιος «ведущий двойной образ жизни, земноводный», из ἀμφί «кругом, вокруг; с обеих или со всех сторон» + βίος «жизнь», далее из праиндоевр. *gweie- «жить». Заимствовано в XVIII веке . Слово «земноводные» происходит от прилагательного «земноводный», далее от соединения слов «земной» + «водный». В разговорной речи животных из отряда бесхвостых земноводных называют « лягушками » или «жабами» (возможно провести параллель с латинскими терминами, используя в качестве основы лат. rana для слова «лягушка» и лат. bufo для слова «жаба»). При попытке наложить разговорные термины «лягушка» и «жаба» на принятую классификацию отряда бесхвостых оказывается, что практически все семейства бесхвостых включают и тех и других. Визуальное отличие одних от других состоит в том, что кожа жаб бородавчатая .
Размеры представителей группы колеблются в широких пределах. Самым большим ныне живущим земноводным считается китайская исполинская саламандра , крупнейшая измеренная особь которой достигала 180 см в длину при массе 59 кг . В прошлом существовали амфибии значительно больших размеров: абсолютный рекорд принадлежит прионозуху — ранне пермскому темноспондилу , длина наиболее крупного экземпляра которого оценивается в более чем 5,5 м .
Самая крупная современная лягушка — африканская лягушка-голиаф , которая может достигать 33 см в длину и весить до 3,3 кг . Весьма крупной также является африканская роющая лягушка , достигающая длины до 24,5 см и массы в 1,4 кг . Самое маленькое земноводное и одновременно самое маленькое позвоночное животное в мире — это лягушка из Папуа—Новой Гвинеи Paedophryne amauensis , впервые описанная в 2012 году — длина её тела оставляет всего 7,7—8 мм . В 2013 году данный вид был включён в список « Десять самых замечательных видов » как мельчайшее из известных позвоночных. Размеры крупных безногих земноводных могут достигать 1,5 м. Так, крупнейший зарегистрированный экземпляр исполинской червяги достигал в длину 152 см. Самые мелкие представители этого отряда относятся к видам крохотная червяга и . Взрослые представители этих видов достигают в длину до 10,4 см и 11,2 см соответственно .


.png)


Размножение и эмбриональное развитие у большинства видов земноводных происходят в водной среде, а взрослые особи обитают на суше. В этом отношении земноводные занимают промежуточное положение между водными ( рыбы ) и наземными позвоночными ( амниоты , то есть рептилии и синапсиды ). Тем не менее, утверждение о том, что современные земноводные «примитивны», является неточным, поскольку их предки претерпели большое количество эволюционных изменений после того, как они разошлись с предками амниот .
Предками наземных позвоночных были лопастепёрые рыбы , которые жили в девонском периоде и принадлежали к группе тетраподоморф (в которую включают и самих четвероногих) . У этих древних морских обитателей развились многосуставчатые, похожие на ноги плавники с пальцами, которые позволяли им ползать по морскому дну. У некоторых рыб развились примитивные лёгкие, которые помогали им дышать воздухом. Они также могли использовать свои сильные плавники, чтобы подняться из воды на сушу, если того потребуют обстоятельства. В конце концов, их костные плавники стали превращаться в конечности, тем самым положив начало жизни на суше. Несмотря на способность передвигаться по суше, многие из тетраподоморф по-прежнему проводили большую часть своей жизни в воде. У них начали развиваться лёгкие, похожие на лёгкие современных земноводных, но они все ещё дышали преимущественно жабрами .

Выраженными переходными признаками обладала лопастепёрая рыба тиктаалик , которая, как предполагается, могла передвигаться по дну на мелководьях рек и озёр или по грязи возле берега . От близких к тиктаалику рыб произошли стегоцефалы, в том числе и лабиринтодонты (в эволюционной таксономии эти примитивные формы относят к земноводным , а в кладистике — к стволовой группе четвероногих ) . Древнейшие скелетные остатки стегоцефалов датируются верхним девоном, возрастом примерно в 372 миллиона лет . Ихтиостега и близкие к ней примитивные стегоцефалы имели четыре крепких конечности, шею, хвост с плавником и череп, очень похожий на череп тетраподоморфных рыб типа Eusthenopteron и Osteolepis , но отличавшийся меньшим кинетизмом . Кожа земноводных стала способной удерживать жидкость организма и сопротивляться высыханию. Кость в подъязычной области за жабрами уменьшилась в размерах и превратилась в стремечко уха земноводных, приспособление, необходимое для слуха на суше . Основное сходство между земноводными и костистыми рыбами заключается в многоскладчатой структуре зубов и парных верхнезатылочных костях, ни один вид не имеет таких особенностей в современном животном мире .
В конце девонского периода (360 миллионов лет назад) моря, реки и озера кишели жизнью, в то время как земля была царством растений и лишена позвоночных , хотя некоторые стегоцефалы (например, ихтиостега) порой выходили на сушу. Считается, что они, возможно, передвигались передними конечностями, волоча заднюю часть подобно тому, как это делают сегодня морские слоны . В начале каменноугольного периода (от 360 до 345 миллионов лет назад) климат стал влажным и тёплым. Образовались обширные пресноводные и морские болота с мхами, папоротниками, хвощами и каламитами. Появились первые настоящие растительноядные членистоногие, тем самым являясь пищей плотоядным амфибиям, которые начали приспосабливаться к земной среде. Других четвероногих на суше не было, и земноводные находились на вершине пищевой цепи . Они были главными наземными хищниками, иногда достигая нескольких метров в длину (например, гигантский прионозух ), охотились на крупных насекомых того периода и на множество видов рыб в воде. Однако развивающимся земноводным по-прежнему нужно было возвращаться в воду, чтобы отложить яйца без скорлупы. До сих пор большинство современных земноводных откладывают яйца в воде или около неё, что отличает их от амниот, которые заменили икру на амниотическое яйцо, заполненное жидкостью и предотвращающее, таким образом, высыхание эмбриона .
После коллапса тропических лесов в каменноугольном периоде амфибии уступили место амниотам, и первые стали исчезать . В триасовый период (от 250 до 200 миллионов лет назад) амниоты продолжали вытеснять земноводных, что привело к уменьшению как размера земноводных, так и их значения в биосфере. Согласно летописи окаменелостей, Lissamphibia , надотряд, который включает в себя всех современных земноводных, которые являются единственными сохранившимися до наших дней, возможно, ответвились от вымерших групп Temnospondyli и Lepospondyli в какой-то период между поздним карбоном и ранним триасом. Относительная нехватка ископаемых свидетельств препятствует точному датированию , однако самое последнее молекулярное исследование, основанное на многолокусном типировании последовательностей, предполагает позднекаменноугольное или раннепермское происхождение современных амфибий .



Происхождение и эволюционные отношения между тремя основными группами амфибий являются предметом споров. Молекулярная филогения 2005 года, основанная на анализе р ДНК , предполагает, что саламандры и червяги более тесно связаны друг с другом, чем с лягушками. Также представляется, что расхождение трёх групп произошло в палеозое или раннем мезозое (около 250 миллионов лет назад), до распада суперконтинента Пангеи и вскоре после их расхождения с лопастепёрыми рыбами. Краткость этого периода и скорость, с которой произошло разделение, могут помочь объяснить относительную редкость окаменелостей примитивных амфибий . Существуют большие пробелы в летописи окаменелостей. Ископаемые остатки Vieraella herbsti были обнаружены в 1961 году в Аргентине и датируются возрастом около 175 миллионов лет (на рубеже ааленского и тоарского ярусов юрского периода ). Строение тела Vieraella близко к современным бесхвостым, и она считается (наряду с чуть более древним Prosalirus bitis ) одним из самых древних их представителей, полностью адаптированным для передвижения прыжками . Некоторые исследователи считают Vieraella самой древней из настоящих лягушек и датируют её возрастом 188—213 млн лет . Обнаружение геробатрахуса ( Gerobatrachus hottoni ) из ранней перми (290 млн лет) в Техасе в 2008 году также дало недостающее звено со многими характеристиками современных лягушек . Он был отнесён к темноспондильным амфибиям, однако обладал признаками, присущими бесхвостым и хвостатым земноводным. Данная находка, по мнению описавших её исследователей, заполнила отсутствующее звено между мезозойскими бесхвостыми и хвостатыми земноводными и их палеозойскими предками. Она также свидетельствовала, что хвостатые и бесхвостые земноводные ближе друг к другу, чем к безногим земноводным . Молекулярный анализ показывает, что расхождение между лягушкой и саламандрой произошло значительно раньше, чем указывают палеонтологические данные. Одно исследование предположило, что последний общий предок всех современных амфибий жил около 315 миллионов лет назад, и что стереоспондильные темноспондильные являются ближайшими родственниками червяг. Тем не менее, большинство исследований поддерживают версию монофилетического происхождения всех современных земноводных в пределах диссорофоидных темноспондилов .
Протолягушки Czatkobatrachus polonicus и Triadobatrachus massinoti известны из нижнего триаса Польши и Мадагаскара соответственно . Хотя известно несколько более древних окаменелостей протолягушек, древнейшей «истинной лягушкой» считается Prosalirus bitis , из отложений в Аризоне , США . Анатомически она сходна с современными лягушками . Древнейшая известная ископаемая червяга Eocaecilia micropodia также найдена в нижней юре Аризоны . Одни из древнейших стволовых хвостатых земноводных — представители семейства из средней и верхней юры Центральной Азии и Западной Европы . Древнейшей известной ископаемой саламандрой является Beiyanerpeton jianpingensis из верхнеюрских отложений северо-восточного Китая .
По мере того, как земноводные эволюционировали из рыб, они приобретали приспособления для жизни на суше. Во время своей жизнедеятельности в воде они пользовались хвостом, как основным органом, отвечающим за их передвижение в воде. Их позвоночник, конечности, пояса конечностей и мускулатура должны были быть достаточно сильными, чтобы отрывать их от земли для передвижения и кормления на суше. Также земноводным нужно было справиться с колебаниями температуры окружающей среды. Их кожа подвергалась воздействию лучей опасного ультрафиолета , которые при водной жизни поглощались самой водой. Поэтому кожа амфибий изменилась, чтобы защищать организм от солнца днём и предотвращать потерю жизненно необходимой жидкости .
Слово «амфибия» происходит от др.-греч. ἀμφί [ amphi ] и βιος [ bios ], что означает «оба вида жизни». Разные интерпретации класса Amphibia присутствуют во всех изданиях работы Карла Линнея « Система природы ». В десятом издании (1758 год), принимаемом за исходный пункт зоологической номенклатуры , Линней относит к амфибиям не только таксоны , ныне известные как беспанцирные , но и пресмыкающихся , некоторых лучепёрых рыб ( Acipenser , Lophius ), хрящевых рыб ( Squalus , Raja , Chimaera ) и бесчелюстных ( Petromyzon ) . В дальнейшем из состава класса были исключены таксоны, не принадлежащие к четвероногим , но при этом в него был включён ряд вымерших представителей . Земноводные в самом широком смысле ( sensu lato ) были разделены на три подкласса, два из которых являются вымершими :
Фактическое количество выделяемых в каждой группе видов зависит от используемой классификации. Двумя наиболее распространёнными системами являются: классификация, принятая на веб-сайте Калифорнийского университета в Беркли , и классификация герпетолога Даррела Фроста и Американского музея естественной истории , доступная как онлайн база данных . Согласно обеим классификациям, всего известно около 8,5 тысяч современных видов земноводных, из которых примерно 90 % составляют лягушки . Точное количество видов и семейств в классификации выше указано по данным Amphibian Species of the World от 31 октября 2022 года .

Классификации земноводных отличаются в зависимости от предпочитаемых их авторами направлений биологической систематики и способов определения клад . В эволюционной таксономии земноводные считаются группой позвоночных, промежуточной между костными рыбами и пресмыкающимися . Под такое определение попадают в основном наземные животные, которые в большинстве своём откладывают икру в воду и развиваются через водную личиночную стадию . Во-первых, это беспанцирные (Lissamphibia) — представители группы, включающей современных земноводных (лягушек, саламандр и червяг) и всех вымерших потомков их ближайшего общего предка . Во-вторых, в соответствии с данным определением, земноводными являются неоднородные формы, которые жили с позднего девона по ранний мезозой и традиционно разделяются на подклассы лабиринтодонтов (Labyrinthodontia) и тонкопозвонковых (Lepospondyli) .
_of_the_extant_amphibians.png)
Если общий предок беспанцирных и амниот включён в класс земноводных, то земноводные оказываются парафилетической группой наряду с рыбами , ящерицами и т. п. Сторонники кладистики , которые не признают не- монофилетические таксоны, считают необходимым переопределить земноводных таким образом, чтобы к ним относились кроновая группа беспанцирных и все вымершие таксоны, более близкие к беспанцирным, чем к амниотам. и соавторы (2020) дали следующее определение Amphibia, соответствующее правилам « »: наибольшая тотальная клада , включающая ( ) , большого сирена ( Siren lacertina ), японскую исполинскую саламандру ( Andrias japonicus ), европейского протея ( Proteus anguinus ) и травяную лягушку ( Rana temporaria ), но не человека разумного ( Homo sapiens ) . В зависимости от принимаемой топологии филогенетического анализа, клада Amphibia в понимании Лорина и коллег включает беспанцирных и парафилетических по отношению к ним темноспондильных (традиционно считаются лабиринтодонтами ) или тонкопозвонковых . Некоторые авторы включают беспанцирных в состав темноспондильных, чтобы избежать парафилии последнего таксона . В классификации группа, включающая беспанцирных и все таксоны, более близкие к ним, чем к амниотам, называется .
В кладистике традиционный подкласс лабиринтодонтов рассматривается как парафилетическая группа, не имеющая уникальных определяющих признаков. То же самое касается и группы стегоцефалов (Stegocephali, или Stegocephalia), которая примерно эквивалента лабиринтодонтам по своему составу. В филогенетической номенклатуре, предложенной Лорином (2020), клада Stegocephali включает все таксоны тетраподоморф, более продвинутые чем тиктаалик , в том числе и Labyrinthodontia, куда, помимо базальной группы , относятся кроновые Tetrapoda. В данной классификации земноводные (как и все четвероногие) находятся в составе стегоцефалов и лабиринтодонтов, а не наоборот .
Все современные амфибии включаются в подкласс беспанцирных, который обычно считается кладой — группой видов, произошедших от общего предка. Однако было высказано предположение, что бесхвостые и хвостатые земноводные произошли от темноспондильных предков, а червяги — от тонкопозвонковых . Беспанцирные традиционно подразделяются на три современных отряда: безногие (Apoda, или Gymnophiona), хвостатые (Caudata, или Urodela) и бесхвостые (Anura, или Salientia). В филогенетической номенклатуре названия Apoda , Caudata и Anura закрепляются за кроновыми группами, а Gymnophiona , Urodela и (прыгающие ) — за тотальными . Также в подкласс включают вымершее семейство саламандроподобных земноводных , иногда выделяемое в собственный отряд .
Некоторые виды земноводных формируют межвидовые гибриды . Так, например, съедобная лягушка представляет собой естественный гибрид прудовой ( Pelophylax lessonae ) и озёрной ( P. ridibundus ) лягушек .
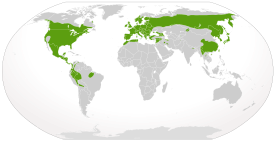
Земноводные распространены на всех континентах, кроме Антарктиды . Исключения составляют большие песчаные пустыни Сахара и Руб-Эль-Хали , наиболее холодные области — Гренландия , Таймыр и другие высокоширотные районы Арктики , Антарктида, а также некоторые острова , удалённые от континентов . Кроме того, естественный ареал земноводных не включал южный остров Новой Зеландии , но после нескольких попыток интродукции по крайней мере два вида ( разноцветная литория и бурая литория ) основали на нём устойчивые популяции . Жаб-аг завезли за пределы естественного ареала в Океанию для уничтожения вредителей. Многие виды имеют ограниченное распространение из-за климатических или географических преград, например, проливов , горных хребтов , пустынь; популяции могут быть изолированы также из-за препятствий, созданных человеком — автотрасс, лесных просек и т. п. В тропических регионах видовое разнообразие, как правило, выше, чем в районах с умеренным климатом . Некоторые виды земноводных адаптированы для выживания в негостеприимных условиях, например, в пустынях или в холодном климате . Так, лесная лягушка , чей ареал частично лежит за северным полярным кругом , на зиму закапывается в землю. Несмотря на глубокое промерзание почвы, высокая концентрация глюкозы в тканях позволяет этой лягушке пережить зиму в состоянии анабиоза .
Из-за водопроницаемой кожи большинство амфибий неспособны жить в солёных и солоноватых водоемах. Единственное исключение — крабоядная лягушка , обитающая в мангровых зарослях Юго-Восточной Азии. Благодаря высокому содержанию мочевины в крови эта лягушка и её головастики могут переносить океаническую солёность (в течение короткого времени) и длительное время жить в солоноватой воде. В солоноватой воде иногда встречается также жаба-ага .

.jpg)
Все земноводные имеют гладкую и тонкую кожу , сравнительно легко проницаемую для жидкостей и газов. Через неё происходит газообмен , что позволяет взрослым амфибиям дышать, не поднимаясь на поверхность воды, а также впадать в спячку на дне водоёмов. Чтобы увлажнять тонкую и нежную кожу, у земноводных развились слизистые железы, в основном на голове, спине и хвосте. Выделение слизи помогает сохранять кожу увлажнённой. Кроме того, у большинства видов амфибий имеются зернистые железы, выделяющие неприятные на вкус или ядовитые вещества. Некоторые токсины земноводных могут быть смертельными для человека, в то время как другие не наносят большого вреда. Основные вырабатывающие яд железы, паротиды , вырабатывают нейротоксины , такие как буфотоксин , и располагаются за ушами у жаб, вдоль спины у лягушек, за глазами у хвостатых и на спине у безногих амфибий .
Как и других позвоночных животных, в коже земноводных выделяется многослойный эпидермис и собственно кожа ( дерма , или кориум). Кожа богата железами , выделяющими слизь . У некоторых слизь может быть ядовитой или облегчающей газообмен. Кожа является дополнительным органом газообмена и снабжена густой сетью капилляров . Роговые образования очень редки, также редки и окостенения кожи: у седлоносной жабы и у рогатых лягушек , например бразильской рогатки , есть костяная пластинка в коже спины, у безногих земноводных имеются чешуйки ; у жаб в коже с возрастом иногда отлагается известь .

Эпидермис состоит из 2—3 слоёв клеток у личинок и 5—7 слоёв у взрослых особей. Клетки наиболее глубокого базального слоя непрерывно делятся и обеспечивают возобновление кожи. У большинства взрослых в ходе метаморфоза земноводных появляется роговой слой , клетки которого уплощаются и ороговевают . Роговой слой обеспечивает защиту от механических повреждений . Роговой слой периодически обновляется в процессе линьки , контролируемой гипофизом и щитовидной железой . У некоторых представителей, например жаб, на коже имеются местные утолщения (часто называемые бородавками). Наружная часть кожи сбрасывается в основном целиком, в отличие от млекопитающих и птиц, у которых она отслаивается хлопьями. Отслоившуюся кожу земноводные часто поедают . В более толстой дерме залегают слизистые и зернистые железы, пигментные клетки, а также кровеносные сосуды и нервы .
Цвет кожи земноводных определяется тремя слоями пигментных клеток, называемых хроматофорами . Эти три слоя клеток состоят из меланофоров (занимающих самый глубокий слой), гуанофоров (образующих промежуточный слой и содержащих много гранул, дающих сине-зелёный цвет) и липофоров (жёлтый, самый поверхностный слой). Изменение цвета, наблюдаемое у многих видов, инициируется гормонами, выделяемыми гипофизом. В отличие от костистых рыб, нервная система не контролирует пигментные клетки напрямую, в результате чего изменение цвета происходит медленнее, чем у рыб. Ярко окрашенная кожа обычно указывает на то, что этот вид ядовит, и цвет кожных покровов является предупреждающим знаком для хищников .
Земноводные имеют скелетную систему, которая структурно идентична другим четвероногим, хотя и с рядом вариаций. Все амфибии имеют четыре конечности, за исключением безногих червяг и нескольких видов саламандр с редуцированными конечностями или без них. Кости полые и лёгкие. Опорно-двигательный аппарат крепкий, чтобы поддерживать голову и тело. Кости полностью окостеневшие, а позвонки сцеплены друг с другом посредством перекрывающихся отростков. Грудной пояс поддерживается мышцами, а хорошо развитый тазовый пояс прикрепляется к позвоночнику парой крестцовых рёбер. Подвздошная кость наклонена вперёд, и тело держится ближе к земле, чем у млекопитающих .
Череп земноводных представляет собой единую малоподвижную структуру . Из-за редукции покровных окостенений , у современных земноводных череп во взрослом состоянии сохраняет много хряща . Сам череп широкий и уплощённый, что повышает объём ротоглоточной полости и увеличивает эффективность всасывания воздуха в неё . Осевой череп платибазальный, то есть имеет широкое основание, позволяющее мозгу располагаться между глазницами . В затылочном отделе окостенению подвергаются лишь парные боковые затылочные кости, каждая из которых несёт по одному затылочному мыщелку , соединяющиеся с шейным позвонком .
Тело разделено на голову, туловище, хвост (у хвостатых) и пятипалые конечности. Голова подвижно соединена с туловищем.
Скелет разделён на отделы:
В позвоночнике выделяют 4 отдела: шейный, туловищный, крестцовый и хвостовой. Число позвонков — от 7 у бесхвостых до 200 у безногих земноводных . У бесхвостых позвонки хвостового отдела срастаются в одну кость .

Шейный позвонок подвижно причленяется к затылочному отделу черепа (он обеспечивает подвижность головы).
К туловищным позвонкам прикрепляются рёбра (кроме бесхвостых, у которых они отсутствуют). Единственный крестцовый позвонок соединён с тазовым поясом . Скелет конечностей образован скелетом пояса конечностей и скелетом свободных конечностей. Плечевой пояс лежит в толще мускулатуры и включает парные лопатки, ключицы и вороньи кости, соединённые с грудиной. Скелет передней конечности состоит из плеча (плечевая кость), предплечья (лучевая и локтевая кости) и кисти (кости запястья, пястья и фаланги пальцев). Тазовый пояс состоит из парных подвздошных, седалищных и лобковых костей, сросшихся между собой. Он прикреплён к крестцовому позвонку через подвздошные кости. В состав скелета задней конечности входят бедро, голень (большая и малая берцовая кости) и стопа (кости предплюсны, плюсны и фаланги пальцев). У бесхвостых кости предплечья и голени сливаются. Все кости задней конечности сильно удлинены, образуя мощные рычаги для передвижения земноводных прыжками . Парные конечности земноводных, в отличие от парных плавников рыб, имеют только внутренний скелет и построены по принципу системы рычагов, соединённых суставами. Связь конечностей с туловищем осуществляется поясами конечностей, устроенными иначе, чем у рыб. Так, в поясе передних конечностей количество элементов сокращается, а сам он теряет связь с черепом, в то время как пояс задних конечностей соединяется с крестцовым позвонком и увеличивается в размерах . Обе пары конечностей устроены по одной схеме, общей для всех наземных позвоночных. В них выделяют три отдела :
Мускулатура подразделяется на мускулатуру туловища и конечностей. Туловищная мускулатура сегментирована. Группы специальных мышц обеспечивают сложные движения рычажных конечностей. На голове расположены поднимающие и опускающие мышцы. У лягушек, например, мышцы лучше всего развиты в области челюстей и конечностей. У хвостатых земноводных также сильно развиты хвостовые мышцы .
Мышечная система амфибий под влиянием наземного образа жизни претерпела значительные изменения. Однообразно построенные сегменты мускулатуры рыб преобразованы в дифференцированные мышцы конечностей, головы, ротовой полости .
Органами дыхания у земноводных являются:
В процессе эволюции земноводные получили большее разнообразие органов для газообмена (внешние жабры, внутренние жабры, лёгкие, кожа), чем любой другой класс позвоночных .

У большинства видов (кроме безлёгочных саламандр и лягушек Barbourula kalimantanensis ) имеются лёгкие не очень большого объёма, в виде тонкостенных мешков, оплетённых густой сетью кровеносных сосудов. Каждое лёгкое открывается самостоятельным отверстием в гортанно-трахейную впадину (здесь расположены голосовые связки, открывающиеся щелью в ротоглоточную полость). Дыхание происходит за счёт изменения объёма ротоглоточной полости: воздух поступает в ротоглоточную полость через ноздри при опускании её дна. При поднимании дна полости воздух проталкивается в лёгкие. У жаб, приспособленных к обитанию в более засушливой среде, кожа ороговевает, и дыхание осуществляется преимущественно лёгкими .

Личинки земноводных обычно обитают в воде, поэтому для дыхания они используют жабры. Они осуществляют газообмен, забирая кислород из омывающей их воды, которая поступает в организм через рот или ноздри, нагнетаемый горловыми движениями. Жабры есть и у животных, у которых нет личинок, плавающих в воде. Тихоокеанская саламандра , относящаяся к лёгочным саламандрам, живородяща и имеет большие мясистые жабры . Обычно жабры исчезают при метаморфозе. Исключение составляют американские протеи рода Necturus . Также среди видов, представители которых обычно теряют жабры, есть неотенические особи, сохраняющие их. Многие представители отряда хвостатых имеют жабры .

Лёгкие земноводных примитивны по сравнению с лёгкими амниот , они имеют несколько внутренних перегородок и большие альвеолы и, следовательно, имеют сравнительно низкую скорость диффузии кислорода, поступающего в кровь . Обмен кислородом осуществляется за счёт . У взрослых дыхание происходит в результате действия насоса: животное сначала всасывает воздух в ротоглоточную полость через наружные ноздри. Вдыхаемый воздух заполняет рот и горло до того, как предыдущий вдох покидает лёгкие, опуская горло с закрытой голосовой щелью. Только тогда воздух из лёгких покидает тело амфибии через ноздри. Амфибия сужает или закрывает наружные ноздри. Мышцы горла сокращаются. Воздух изо рта и глотки поступает в лёгкие через открытую голосовую щель. Голосовая щель снова закрывается, воздух остаётся вне рта и глотки. Механизм последнего отличается от вдоха: воздух выталкивается гибкими лёгкими и движениями стенки туловища. Однако последние происходят реже, чем движения горла . Лёгкие земноводных кажутся примитивными по сравнению с амниотами. Их внутренняя поверхность образует сложные складки. Между ними находятся камеры, то есть лёгочные пузыри, где происходит газообмен. Несколько альвеолярных перегородок разделяют большие альвеолы (иногда их вообще не считают альвеолами). Размер лёгочных мешков лягушки составляет 150—600 мкм. Они выстланы эпителием , образованным клетками одного типа (некоторые авторы различают два типа клеток, покрытым слоем сурфактанта .
Однако большинство земноводных способно производить газообмен с водой или воздухом через кожу. Чтобы обеспечить достаточное кожное дыхание, поверхность кожи должна оставаться влажной, чтобы кислород мог диффундировать с достаточно высокой скоростью. Поскольку концентрация кислорода в воде увеличивается как при низких температурах, так и при высоких скоростях потока, водные амфибии в этих ситуациях могут полагаться в первую очередь на кожное дыхание. Титикакский свистун может всю жизнь дышать кожей. Многие водные саламандры и все головастики имеют жабры на личиночной стадии, а некоторые (например, аксолотль ) имеют жабры всю жизнь или во взрослом состоянии . Большинство земноводных способны осуществлять газообмен через кожу как в воде, так и на воздухе .

Кровеносная система замкнутая, сердце трёхкамерное со смешиванием крови в желудочке (кроме безлёгочных саламандр, которые имеют двухкамерное сердце). Температура тела зависит от температуры окружающей среды (являются холоднокровными животными) .
Кровеносная система состоит из большого и малого (лёгочного) кругов кровообращения. Появление второго круга связано с приобретением лёгочного дыхания. Сердце состоит из двух предсердий (в правом предсердии кровь смешанная, преимущественно венозная, а в левом — артериальная) и одного желудочка. Внутри стенки желудочка образуют складки, препятствующие смешиванию артериальной и венозной крови. Из желудочка выходит артериальный конус, снабжённый спиральным клапаном .
Артерии :
Малый круг — лёгочный, начинается кожно-лёгочными артериями, несущими кровь к органам дыхания (лёгким и коже); от лёгких обогащённая кислородом кровь собирается в парные лёгочные вены, впадающие в левое предсердие. Большой круг кровообращения начинается дугами аорты и сонными артериями, которые ветвятся в органах и тканях. Венозная кровь по парным передним полым венам и непарной задней полой вене попадает в правое предсердие. Кроме того, в передние полые вены попадает окисленная кровь от кожи, и поэтому кровь в правом предсердии смешанная. Помимо обычного сердца у всех земноводных есть лимфатические сердца , которые расположены перед местами впадения в вены лимфатических сосудов .
У земноводных есть ювенильная стадия и взрослая стадия, и системы кровообращения у них разные. На стадии молоди (или головастика) кровообращение похоже на кровообращение рыбы; двухкамерное сердце перекачивает кровь через жабры, где она насыщается кислородом, и распространяется по телу и обратно к сердцу по единой петле. На взрослой стадии земноводные (особенно лягушки) теряют жабры и у них развиваются лёгкие. Когда желудочек начинает сокращаться, деоксигенированная (наполненная кислородом) кровь перекачивается через лёгочную артерию в лёгкие. Затем продолжающееся сокращение перекачивает насыщенную кислородом кровь по всему телу .
Практически все земноводные питаются только подвижной добычей. На дне ротоглоточной полости находится мускулистый язык . У бесхвостых он передним концом прикрепляется к нижним челюстям, при ловле мелкой добычи, например насекомых, язык выбрасывается изо рта, к нему прилипает добыча; длина языка у самых крупных достигает 7-11 сантиметров. Большинство амфибий проглатывают свою добычу целиком, не пережёвывая её, потому что у них объёмный желудок. Короткий пищевод выстлан крохотными отростками, которые помогают продвигать пищу в желудок, а слизь, вырабатываемая железами рта и глотки, облегчает её прохождение. На челюстях имеются зубы, служащие только для удержания добычи. У лягушек они расположены только на верхней челюсти, а у представителей семейства настоящие жабы они вообще отсутствуют . Единственным видом бесхвостых, имеющим зубы на обоих челюстях является полнозубая квакша . Доказано, что нижние зубы отсутствовали уже у предка всех современных лягушек около 230 млн лет назад, и возникли заново у этого вида 5-17 млн лет назад. Таким образом, данный вид является примером нарушения закона Долло .
В ротоглоточную полость открываются протоки слюнных желёз, секрет которых не содержит пищеварительных ферментов. Из ротоглоточной полости пища по пищеводу поступает в желудок, оттуда в двенадцатиперстную кишку . Сюда открываются протоки печени и поджелудочной железы . Переваривание пищи происходит в желудке и в двенадцатиперстной кишке. Тонкий кишечник переходит в прямую кишку , которая образует расширение — клоаку . Так же имеется жёлчный пузырь, накапливающий в себе желчь, выработанную печенью .

Земноводные обладают поджелудочной железой, печенью и жёлчным пузырём. Печень обычно крупная. Её размер определяется её функцией в качестве места хранения гликогена и жира и может меняться в зависимости от времени года по мере того, как эти запасы накапливаются или расходуются. Жировая ткань является ещё одним важным средством сохранения источника энергии, и это происходит в брюшной полости (во внутренних структурах, называемых жировыми телами), под кожей и, у некоторых саламандр, в хвосте .
Органы выделения — парные туловищные почки , от которых отходят мочеточники , открывающиеся в клоаку. В стенке клоаки имеется отверстие мочевого пузыря , в который стекает моча, попавшая в клоаку из мочеточников. В туловищных почках не происходит обратного всасывания воды. После наполнения мочевого пузыря и сокращения мышц его стенок концентрированная моча выводится в клоаку и выбрасывается наружу. Своеобразная сложность такого механизма объясняется необходимостью земноводных сохранять большее количество влаги. Поэтому моча не удаляется сразу из клоаки, а попав в неё, предварительно направляется в мочевой пузырь. Часть продуктов обмена и большое количество влаги выделяется через кожу .
Две почки расположены дорсально полости тела. Их работа состоит в том, чтобы фильтровать кровь от метаболических отходов и транспортировать мочу через мочеточники в мочевой пузырь, где она хранится, прежде чем периодически выходить через клоакальное отверстие. Личинки и большинство взрослых водных амфибий выделяют азот в виде аммиака в больших количествах разбавленной мочи, в то время как наземные виды, нуждающиеся в большей экономии воды, выделяют менее токсичный продукт, мочевину. Некоторые древесные лягушки с ограниченным доступом к воде выделяют большую часть отходов своего метаболизма в виде мочевой кислоты .
Личинки и большинство взрослых амфибий, ведущих водный образ жизни, выделяют азот в виде аммиака, растворённого в большом количестве разбавленной мочи. Амфибия способна вырабатывать такое большое количество мочи благодаря значительной скорости клубочковой фильтрации (её снижение — один из первых эффектов обезвоживания амфибии) и высокой реабсорбционной способности канальцев. С другой стороны, наземные виды, у которых скорость клубочковой фильтрации даже выше, чем у водных видов, вынуждены в большей степени экономить воду, выделяя менее токсичную мочевину. Этот парадокс объясняется тем, что, в отличие от водных амфибий, они могут в значительной степени полагаться на почки для выделения продуктов азотистого обмена. Водные земноводные в основном используют для этого кожу. Кроме того, почечные канальцы также могут реабсорбировать воду, что обусловлено действием антидиуретического гормона (хотя и в гораздо меньшей степени, чем у млекопитающих) .
Амфибии, ведущие пресноводный образ жизни, в качестве продуктов азотистого обмена выделяют аммиак (аммониотеличный обмен), а наземные формы — мочевину (уреотеличный обмен) .
Эти особенности не позволили земноводным полностью перейти к наземному образу жизни .
В сравнении с рыбами вес головного мозга земноводных больше. Вес головного мозга в процентах от массы тела составляет у современных хрящевых рыб 0,06—0,44 %, у костных рыб 0,02—0,94, у хвостатых земноводных 0,29—0,36, у бесхвостых 0,50—0,73 % .

Головной мозг земноводных относительно прост, но структурно такой же, как у рептилий, птиц и млекопитающих. Их мозг удлинён, за исключением червяг, и содержит обычные двигательные и сенсорные области тетраподоморфов . Считается, что шишковидное тело вырабатывает гормоны, участвующие в спячке .
Состоит из 5 отделов :
Передний мозг, даже без обонятельных луковиц, у представителей семейств настоящие лягушки и жабы составляет 40 % массы мозга, у хвостатых и того больше — у видов рода Амбистомы 56 % . Передний мозг состоит из двух полушарий. Эти полушария делятся на дорсальную и вентральную области, а те, в свою очередь, на латеральную и медиальную. Латеральная и медиальная области делятся на дорсальные, латеральные и медиальные .
Головной мозг посылает сигналы всему телу в процессе передвижения земноводного. От головного мозга отходят 10 пар черепных нервов (I—X) . I (обонятельный) и II (зрительный) нервы передают раздражения, полученные соответствующими органами чувств. Нервы III, IV и VI обеспечивают двигательную иннервацию глазного яблока. Большой V (тройничный) нерв несет как чувствительные, так и двигательные волокна, как и малый VII (лицевой) нерв. VIII (статоакустический) нерв снова связан с соответствующим органом чувств, кожей и глазами, как и нервы, обеспечивающие функционирование боковой линии. Нервы IX и X (порочные) соответствуют определенным группам нервных волокон, начинающихся от средне-каудальной части продолговатого мозга и идущих главным образом к гортани и языку. Рядом с началом волокон X нерва из ядра в шейном отделе спинного мозга выходит также XI нерв. По мнению некоторых специалистов, XII (двенадцатого подъязычного) нерва не существует, хотя его ядро имеется. Соответствующие нервные волокна иногда относят к первому спинномозговому нерву .
.jpg)
Глаза приспособлены к функционированию в воздушной среде. У земноводных глаза похожи на глаза рыб, однако не имеют серебристой и отражательной оболочек, а также серповидного отростка . Недоразвиты глаза только у протеев . У высших земноводных есть верхние (кожистые) и нижние (прозрачные) подвижные веки . Мигательная перепонка (вместо нижнего века у большей части бесхвостых) выполняет защитную функцию. Слёзные железы отсутствуют, но есть гардерова железа , секрет которой смачивает роговицу и предохраняет её от высыхания. Роговица выпуклая. Хрусталик имеет форму двояковыпуклой линзы, диаметр которой меняется в зависимости от освещения; аккомодация происходит за счёт изменения расстояния хрусталика до сетчатки. У многих развито цветное зрение. Глаза головастиков лишены век, но при метаморфозе роговица становится более куполообразной, хрусталик уплощается, развиваются веки и связанные с ними железы и протоки. Глаза взрослых являются усовершенствованием глаз беспозвоночных и были первым шагом в развитии более продвинутых глаз позвоночных. Они обеспечивают цветовое зрение и глубину резкости. В сетчатке находятся зелёные палочки, восприимчивые к широкому диапазону длин волн .
Глаза бесхвостых расположены в верхней части головы и часто выдвинуты вперёд. Это обеспечивает бесхвостым широкое поле зрения; лягушка может полностью погрузиться в воду, оставив над поверхностью лишь глаза. Глаза защищены подвижными веками и дополнительно прозрачной неподвижной мембраной, предохраняющей глаза во время пребывания в воде. Цвет радужной оболочки и форма зрачка у разных видов отличаются .

Бесхвостые лучше видят дальние объекты, чем ближние. Квакающие лягушки немедленно умолкают при виде потенциальной угрозы и даже её тени, но чем ближе объект, тем хуже они его видят . Когда лягушка выстреливает языком в сторону добычи, она реагирует на маленький движущийся объект, который плохо различим. Она прицеливается заранее, так как особенности анатомии вынуждают её закрывать глаза при вытягивании языка . Вопрос о наличии у бесхвостых цветного зрения не решён. Опытами была доказана положительная реакция бесхвостых на голубой свет .
Лягушки обладают уникальным среди позвоночных зрительным аппаратом. В ходе исследований было обнаружено, что почти 95 % информации поступает в рефлекторный отдел мозга. Это приводит к тому, что лягушка не видит, где она находится. Основной вывод — лягушки видят только движущиеся предметы .
Органы обоняния функционируют только в воздушной среде, представлены парными обонятельными мешками. Их стенки выстланы обонятельным эпителием. Открываются наружу ноздрями , а в ротоглоточною полость — хоанами .
_-_Teichmolch_(30787893742).jpg)
В органе слуха новый отдел — среднее ухо . Наружное слуховое отверстие закрывает барабанная перепонка , соединённая со слуховой косточкой — стремечком . Стремечко упирается в овальное окно, ведущее в полость внутреннего уха, передавая ему колебания барабанной перепонки. Для выравнивания давления по обе стороны барабанной перепонки полость среднего уха соединена с ротоглоточной полостью слуховой трубой. Некоторые червяги обладают электрорецепторами, которые позволяют им находить предметы вокруг себя, когда они погружены в воду. Уши хорошо развиты у лягушек. Наружного уха нет, но большая круглая барабанная перепонка лежит на поверхности головы сразу за глазом. Она вибрирует под воздействием звука, и звук передаётся далее через стремечко, единственную кость, во внутреннее ухо. Таким образом земноводному слышны только высокочастотные звуки, такие как брачные крики. Существует участок специализированных волосковых клеток во внутреннем ухе, способный обнаруживать более низкочастотные звуки. Другой особенностью, уникальной для лягушек и саламандр, является колумелла-жаберный комплекс, примыкающий к слуховой капсуле. Уши саламандр и червяг менее развиты, чем у лягушек, поскольку они не общаются друг с другом с помощью звука . Как правило, земноводные слышат как на земле, так и под водой. Внешнего уха у них нет, но, как правило , позади каждого глаза присутствует барабанная перепонка. Звук вызывает вибрацию перепонок, которые передают её в среднее и внутреннее ухо. Размер барабанных перепонок и дистанция между ними коррелируют с частотой звука, на которой квакает данная лягушка. У некоторых видов, например у лягушки-быка , размер перепонок относительно размера глаз указывает на половую принадлежность. У самцов перепонки больше глаз, а у самок они схожего размера. Как правило, земноводные не полагаются только на слух, и даже на резкий звук они не будут реагировать, пока не увидят его источник .
Органом осязания является кожа, содержащая осязательные нервные окончания. У водных представителей и головастиков имеются органы боковой линии . Органы вкуса расположены в ротовой полости. Предполагается, что лягушка воспринимает только горькое и солёное .
Земноводные — первая группа четвероногих, у которых развился орган Якобсона . Это парная полость, выстланная обонятельным эпителием. Возможно, он хранит молекулы ароматов, которые проникают в него через носовую полость, получая раздражители даже после того, как запах рассеялся. У представителей отряда бесхвостых, например у серой жабы, имеется отдельная полость, также выстланная обонятельным эпителием, которая соединяется с носовой полостью тонкой трубкой диаметром 18,5 мкм. Это указывает на небольшое количество пахучего вещества, которое может попасть в орган .
Все земноводные раздельнополые. У большинства земноводных оплодотворение наружное (в воде). В период размножения яичники , наполненные зрелыми яйцеклетками , заполняют у самок почти всю брюшную полость. Созревшие икринки выпадают в полость тела, попадают в воронку яйцевода и, пройдя по нему, через клоаку выводятся наружу. Самцы имеют парные семенники . Отходящие от них семявыводящие канальцы попадают в мочеточники, одновременно служащие самцам семяпроводами. Они также открываются в клоаку .
Работа половых желёз земноводных находится под контролем железистого гипофиза , который, в свою очередь, контролируется гипоталамусом . Эти два отдела соединены сосудами. Гипофиз, в свою очередь, влияет на половые железы через эозинофильные и базофильные клетки посредством секреции гонадотропинов. Разрушение гипофиза связано с потерей половых органов, а у самок ещё и с исчезновением вторичных половых признаков .
Семенники, или мужские гонады, осуществляют гаметогенез у самцов. Они описаны как имеющие простую яйцевидную структуру. Это относится к лягушкам и большинству представителей отряда хвостатых. Имеются исключения: у и некоторых лягушек неотропической области они несколько шире и длиннее, как и у Gymnophion . У последних количество долей яичка не меняется в течение жизни земноводного, тогда как у вышеупомянутых хвостатых число долей может увеличиваться со временем .
Семенники содержат созревающие зародышевые клетки, находящихся на разных стадиях развития . Как правило, самец не имеет копулятивного органа . Точно так же женские гонады — яичники — проводят оогенез . Он сходен у бесхвостых амфибий, хвостатых земноводных и червяг, хотя есть и различия (например, у встречаются ооциты с 8-клеточными ядрами, у саламандр встречаются и многоядерные ооциты). Яичники содержат фолликулы, внутри которых находятся созревающие и мейотически делящиеся ооциты .

В жизненном цикле земноводных чётко выделяются четыре стадии развития: яйцо (икринка), личинка ( головастик ), период метаморфоза , взрослая особь .
Большинство амфибий проходят через метаморфоз — процесс значительных морфологических изменений после рождения. При типичном развитии земноводных яйца откладываются в воду, а личинки приспосабливаются к водному образу жизни. Лягушки, жабы и саламандры вылупляются из яйца в виде личинок с наружными жабрами. Метаморфоз у земноводных регулируется концентрацией в крови тироксина, стимулирующего метаморфоз, и пролактина, противодействующего действию тироксина . Конкретные события зависят от пороговых значений для различных тканей. Поскольку большая часть эмбрионального развития происходит вне родительского тела, оно подвержено многим адаптациям из-за конкретных обстоятельств окружающей среды. По этой причине головастики могут иметь роговые гребни вместо зубов, усикоподобные отростки кожи или плавники. Они также используют сенсорный орган боковой линии, подобный органу рыб. После метаморфоза эти органы становятся излишними и будут реабсорбироваться в результате контролируемой гибели клеток, называемой апоптозом. Разнообразие приспособлений к конкретным условиям окружающей среды у амфибий велико, и многие открытия ещё не сделаны .

Земноводные относятся к анамниям . Их яйца лишены зародышевых оболочек, в связи с чем для развития им требуются постоянное увлажнение .

Подавляющее большинство земноводных откладывают икру в пресных водоёмах, однако известны и исключения: червяги откладывают яйца в почву и обвивают место закладки, тем самым увлажняя яйца собственной слизью, , гигантские саламандры , и некоторые другие амфибии делают кладку на суше. Даже в этих случаях яйца нуждаются в повышенной влажности окружающей среды, обеспечение которой ложится на родителя. Известны виды, которые носят икринки на своём теле: самка прикрепляет их к животу, а самцы жаб-повитух обматывают шнурообразную кладку вокруг задних ног. Особенно необычно выглядит забота о потомстве у суринамской пипы — оплодотворённая икра вдавливается самцом в спину самки и последняя носит её на себе, пока из икры не вылупятся молодые пипы. В яйце зародыш подвешен в перивителлиновой жидкости и окружён полупроницаемыми студенистыми капсулами, а желточная масса обеспечивает питательные вещества. Когда личинки вылупляются, капсулы растворяются ферментами, выделяемыми железой на кончике рыла. Яйца некоторых саламандр и лягушек содержат одноклеточные зелёные водоросли. Они проникают в желейную оболочку после откладывания яиц и могут увеличить снабжение эмбриона кислородом посредством фотосинтеза. Похоже, что они ускоряют развитие личинок и снижают смертность. Яйца могут откладываться по одному, несколько или длинными нитями. Места для откладывания яиц: вода, грязь, норы, мусор и растения, а также некоторые амфибии откладывают икру под брёвнами или камнями .
Размер и форма агломерата икры характерна для каждого вида. Лягушки семейства Ranidae имеют тенденцию к шарообразным кластерам, а жабы к длинным цилиндрическим кластерам. Крошечный кубинский свистун откладывает икринки по одной и зарывает их во влажную почву. Leptodactylus pentadactylus создаёт пенное гнездо в норе, откладывая в него около тысячи икринок. Головастики появляются на свет, когда вода заполняет нору, а иногда развитие полностью происходит в гнезде. Красноглазая квакша откладывает икринки на листья, расположенные над поверхностью водоёма. Вылупляясь, головастики падают с листьев в воду. У некоторых видов на определённой стадии развития зародыши в икринках могут уловить вибрации, вызываемые хищниками (осами, змеями), и вылупиться раньше времени, чтобы приобрести мобильность и избежать гибели . В общем, длительность стадии развития зародышей в икре зависит от конкретного вида и окружающих условий. Как правило, головастики вылупливаются в течение недели после того, как капсула икринки распадается под воздействием гормона , вырабатываемого зародышем .


Из икринок вылупляются личинки (у бесхвостых земноводных называемые головастиками ), ведущие водный образ жизни. По своему строению личинки напоминают рыб: у них отсутствуют парные конечности, дышат жабрами (наружными, затем внутренними) и кожей; имеют двухкамерное сердце и один круг кровообращения, органы боковой линии . Некоторые червяги, альпийские саламандры и некоторые африканские живородящие жабы являются живородящими. Их личинки питаются железистым секретом и развиваются в яйцеводе самки, часто в течение длительного времени. Другие земноводные, но не червяги, яйцеживородящие. Яйца сохраняются в теле родителей или на нём, но личинки питаются желтками своих яиц и не получают питания от взрослых особей. Личинки появляются на разных стадиях своего роста, до или после метаморфоза, в зависимости от их вида .

Головастики лягушек, в отличие от других амфибий, не похожи на взрослых особей . Личинки бесхвостых, появляющиеся из икринок, известны как «головастики». Они ведут полностью водный образ жизни, но известно одно исключение — головастики вида Nannophrys ceylonensis полусухопутные и обитают среди мокрых камней . Их тело, как правило, имеет овальную форму, хвост длинный, сплющенный по вертикали, приспособлен для плаванья. У головастиков хрящевой скелет, глаза лишены век, есть боковая линия , для дыхания служат жабры . В начале у головастиков появляются внешние жабры, а позже внутренние (жаберный мешок закрывает жабры и передние ноги). Развивающиеся лёгкие служат дополнительным дыхательным органом. Некоторые виды проходят метаморфоз ещё в икринке, и из икринок вылупляются лягушата. У головастиков нет настоящих зубов, но у многих видов челюсти покрыты параллельными рядами кератиновых зубчиков (два ряда на верхней челюсти, три ряда на нижней и роговой клюв). Количество рядов и точная морфология рта варьируют у разных видов и могут служить диагностическим признаком. Головастики семейства пиповые (за исключением рода Hymenochirus ) имеют пару передних усиков, делающих их похожими на небольших сомиков . Головастики имеют хрящевой скелет, жабры для дыхания (сначала внешние жабры, позже внутренние), систему боковых линий и большие хвосты, которые они используют для плавания . У недавно вылупившихся головастиков вскоре появляются жаберные мешки, закрывающие жабры. Лёгкие развиваются рано и используются как вспомогательные органы дыхания, головастики поднимаются к поверхности воды, чтобы глотать воздух. Некоторые виды завершают своё развитие внутри яйца и вылупляются прямо в маленьких лягушек. У этих личинок нет жабр, но есть специальные участки кожи, через которые происходит дыхание. Хотя у головастиков нет настоящих зубов, у большинства видов челюсти имеют длинные параллельные ряды небольших ороговевших структур, называемых керадонтами, окружённых роговым клювом. Передние ноги формируются под жаберным мешком, а задние становятся видными через несколько дней . Головастики, как правило, травоядны и питаются водорослями, отфильтрованными из воды через жабры. Некоторые виды являются хищниками уже на стадии головастика и питаются насекомыми; головастики ( Osteopilus septentrionalis практикуют каннибализм ), а также поедают мелких рыб. Головастики, рано отрастившие ноги, могут стать жертвой своих собратьев .
На головастиков охотятся рыбы, саламандры , хищные жуки и птицы (например, зимородок ). Некоторые головастики, в том числе личинки Bufo marinus , ядовиты. У разных видов бесхвостых стадия головастика продолжается от недели до нескольких месяцев и зависит от стратегии размножения .
После завершения стадии головастика бесхвостые проходят процесс метаморфоза , во время которого системы организма резко перестраиваются во взрослую форму. При метаморфозе происходят быстрые изменения в организме, так как полностью меняется образ жизни лягушки. Как правило, метаморфоз длится около суток. Он начинается с выработки гормона тироксина , который влияет на развитие тканей. Так, изменение органов дыхания включает в себя развитие лёгких параллельно с исчезновением жабр и жаберного мешка. Передние конечности становятся видимыми. Нижняя челюсть приобретает характерную для хищника форму, кишечник укорачивается. Нервная система адаптируется под стереоскопическое зрение и слух, а также под новые способы передвижения и питания. Глаза сдвигаются выше, формируются веки и связанные с ними железы. Модифицируются органы слуха (появляется слуховая перепонка и среднее ухо). Кожа становится толще и крепче, боковая линия исчезает (у большинства видов), формируются кожные железы. На последнем этапе метаморфоза исчезает хвост, ткани которого идут на развитие конечностей .Спиралевидный рот с роговыми зубными гребнями резорбируется вместе со спиральной кишкой. У животного развивается большая челюсть, и его жабры исчезают вместе с жаберным мешком. Быстро растут глаза и ноги, формируется язык. Есть связанные изменения в нейронных сетях, такие как развитие стереоскопического зрения и потеря системы боковой линии. Через несколько дней хвост реабсорбируется из-за более высокой концентрации тироксина, необходимой для этого .



Типичная личинка хвостатых земноводных, когда вылупляется из икры, больше похожа на взрослую особь, чем у бесхвостых. Перистые наружные жабры прикреплены к хрящевым жаберным дугам. Имеются широкие спинные или хвостовые плавники. Глаза лишены век. На нижней челюсти и на верхней челюсти имеются зубы, три пары перистых наружных жабр, слегка уплощённое тело и длинный хвост со спинным и брюшным плавниками. Передние лапы могут быть частично развиты, а задние в зачаточном состоянии у видов, обитающих в прудах, и могут быть немного лучше развиты у видов, размножающихся в проточных водах. У головастиков, живущих в прудах, часто есть пара стабилизаторов, стержнеобразных структур по обеим сторонам головы, которые не дают жабрам соприкасаться с отложениями. Некоторые представители родов Ambystoma или Dicamptodon производят личинок, которые никогда полностью не созревают, в зависимости от вида и популяции. Они могут постоянно оставаться в личиночном состоянии. Это явление называется неотенией .

У тритонов и саламандр метаморфозы менее интересны, чем у лягушек. Это связано с тем, что личинки уже плотоядны и продолжают питаться как хищники, когда становятся взрослыми, поэтому их пищеварительная система не изменяется. Их лёгкие начинают функционировать рано, работают они довольно плохо, но головастики не дышат ими так активно, как головастики лягушек. Другие метаморфозы, это уменьшение размера или потерю хвостовых плавников, закрытие жаберных щелей, утолщение кожи, развитие век и определённые изменения в зубном ряду и строении языка. Наибольшую уязвимость при трансформации проявляют хвостатые земноводные, так как развиваемая ими скорость плавания значительно падает, а длинный хвост, медленно проходящий различные метаморфозы является препятствием на суше . Трансформация может произойти примерно через 3 месяца . Жабры никогда не закрываются, они рассасываются непосредственно перед выходом из воды. Другие изменения включают уменьшение размера или потерю спинного плавника, закрытие жаберных щелей, утолщение и укрепление кожи, развитие век и некоторые изменения в зубном ряду и строении языка. Взрослые хвостатые весной и летом возвращаются в воду, а зимой живут на суше. Для адаптации к воде необходим гормон пролактин, а адаптация к наземным условиям зависит от тироксина . Наружные жабры, рассосавшиеся до первого выхода на сушу, после возвращения в воду не восстанавливаются .

При вылуплении типичная личинка саламандры имеет глаза без век, зубы как в верхней, так и в нижней челюсти, три пары перистых наружных жабр и длинный хвост со спинным и брюшным плавниками. Передние конечности могут быть частично развиты, а задние конечности могут быть рудиментарными у видов, живущих в пруду, но могут быть более развиты у видов, которые размножаются в движущейся воде. Личинки прудового типа часто имеют пару балансиров, стержнеобразных структур по обе стороны головы, которые могут предотвратить засорение жабр отложениями . Оба они способны размножаться . Саламандры без лёгких из семейства Plethodontidae ведут наземный образ жизни и откладывают небольшое количество непигментированных яиц скоплением среди влажной опавшей листвы. Каждое яйцо имеет большой желточный мешок, и личинка питается им, пока развивается внутри яйца, полностью сформировавшись в молодую саламандру. Самка саламандры часто высиживает яйца. Было замечено, что у представителей рода Ensatinas самка обвивается вокруг них и прижимается к ним областью горла, массируя их слизистым секретом .
У некоторых земноводных есть личинки, которые никогда полностью не развиваются во взрослую форму, это состояние, известное как неотения, присутствует, например, у аксолотля. Неотения возникает, когда скорость роста животного очень низкая, и обычно связана с неблагоприятными условиями, такими как низкая температура воды, которая может изменить реакцию тканей на гормон тироксин, а также нехватка еды. Существует пятнадцать видов неотенических саламандр, в том числе виды американских протеев ( Necturus) , европейский протей ( Proteus) и угревидная амфиума ( Amphiuma ) , а также множество других видов, таких как северо-западная саламандра ( Ambystoma gracile ) и тигровая саламандра ( A. tigrinum ), которые применяют эту стратегию при соответствующих условиях окружающей среды и других обстоятельствах . Они также могут трансформироваться и во взрослое животное. В обоих стадиях животные готовы и способны к размножению. Неотения возникает при очень низкой скорости роста, обычно связанной с неблагоприятными условиями, такими как низкая температура воды, которая может изменить реакцию ткани на гормон тироксин. Другие факторы, которые могут препятствовать метаморфозу, включают нехватку пищи, недостаток микроэлементов и внутривидовую конкуренцию. Амбистома тигровая ( Ambystoma tigrinum ) также иногда проявляет неотению, в этой форме она может достигать действительно больших размеров. Зрелая особь ведёт наземный образ жизни, а личинка обитает в воде, приобретая способность к размножению (сохраняя личиночное состояние) .

Развитие личинок цейлонский рыбозмей из Шри-Ланки хорошо изучено. Личинки, похожие на угрей, вылупляются из яиц и направляются в воду. У них три пары наружных красных перистых жабр, голова с двумя рудиментарными глазами и короткий хвост с плавниками.В основном они активны ночью, вскоре теряют жабры и совершают вылазки на сушу. Метаморфозы постепенные. К десятимесячному возрасту у них развилась заострённая голова с сенсорными щупальцами возле рта, и они потеряли глаза, системы боковых линий и хвосты. Кожа утолщается, тело делится на сегменты. К этому времени они уже вырыли себе нору и живут на суше .
Большинство видов червяг живородящи. Типичным из них является Typhlonectes Compressicauda , вид из Южной Америки. Одновременно в яйцеводе может развиваться до девяти личинок. Они довольно длинные, имеют парные мешкообразные жабры, маленькие глаза и специализированные зубы-скребки. Сначала они питаются желтками яиц, но по мере того, как этот источник питания иссякает, они начинают грызть реснитчатые эпителиальные клетки, выстилающие яйцеводы. Это стимулирует секрецию жидкости, богатой липидами и мукопротеинами, которыми они питаются вместе с соскобами со стенки яйцеводов. Они могут увеличить свою длину в шесть раз и быть в два раза длиннее своей матери до рождения. К этому времени они претерпели метаморфоз, потеряли глаза и жабры, у них развилась более толстая кожа и ротовые щупальца, а зубы снова исчезли и рассосались. Постоянный набор зубов вырастает вскоре после рождения .
Кольчатая червяга ( Siphonops annulatus ) выработала уникальное приспособление для целей размножения. Потомство питается слоем кожи, специально выращенным взрослыми в результате явления, известного как . Выводок питается партиями в течение примерно семи минут с интервалом примерно в три дня, что даёт коже возможность восстановиться. Между тем, было замечено, что они проглатывают жидкость, выделяемую из материнской клоаки .

_(16063246275).jpg)

Процесс метаморфоза у бесхвостых амфибий происходит довольно быстро, тогда как у более примитивных саламандр и безногих земноводных он сильно растянут во времени .
Продолжительность жизни бесхвостых в естественных условиях мало изучена. С использованием методов скелетохронологии продолжительность жизни лягушки была измерена по сезонным изменениям роста фаланг пальцев. Полученные данные позволяют предположить, что максимальная продолжительность жизни взрослой особи составляет 10 лет, а учитывая стадию головастика, которая у этого вида длится около 4 лет, срок жизни этих лягушек равен 14 годам . В неволе зарегистрирована продолжительность жизни бесхвостых до 40 лет, этого возраста достигла обыкновенная жаба . Жаба-ага жила в неволе 24 года, а лягушка-бык — 14 лет . Домашние африканские роющие лягушки могут прожить в неволе до 35 лет .
Саламандры могут жить до 43 лет (огненная, китайская и японская исполинские саламандры) .
Забота о потомстве у амфибий мало изучена, но, как правило, чем больше количество яиц в кладке, тем меньше вероятность проявления какой-либо степени родительской заботы. У 20 % видов земноводных одна или обе взрослые особи играют определённую роль в заботе о потомстве . Те виды, которые размножаются в небольших водоёмах или других специализированных местообитаниях, как правило, заботятся о потомстве и методы помощи у них достаточно интересны .

Многие лесные саламандры откладывают яйца под брёвна или камни на суше. вынашивает яйца и охраняет их от хищников, пока эмбрионы питаются желтками своих яиц. Когда они полностью разовьются, они вырываются из икры, будучи уже полностью сформированными саламандрами. Самцы некоторых видов примитивных саламандр, выкапывают подводное гнездо, где самки откладывают икру. Затем самец охраняет это место в течение двух или трёх месяцев до вылупления яиц .

Южноамериканская лягушка Engystomops pustulosus строит из пены гнездо, в которое она откладывает икру. Пеноматериал состоит из белков и лектинов и, возможно, имеет антибактериальные свойства . Несколько пар лягушек могут строить совместное гнездо. В этом случае вначале строится «плот», а затем лягушки откладывают икру в его центре, чередуя откладывание икры и формирование пеноматериала, завершая процесс созданием пенного слоя поверх икры . Самцы лягушек из семейства филломедуз строят чаши из глины высотой 10, а шириной 30 сантиметров, куда самки откладывают икру. Головастики живут в посудине до превращения в лягушку; так они более защищены от хищников .

Самец крохотного вида защищает скопление яиц, спрятанное под камнем или бревном. После вылупления икры самец переносит на спине головастиков. Самец жабы-повитухи наматывает яичные нити вокруг своих бёдер и носит яйца в течение восьми недель. Он держит их влажными, а когда они готовы вылупиться, посещает пруд или канаву и выпускает головастиков . Сумчатая лягушка откладывает яйца на землю. Когда они вылупляются, самец носит головастиков в выводковых сумках на задних лапах. Самка заботливой лягушки ( Rheobatrachus spp ) выращивала личинок в своём желудке после проглатывания яиц. Головастики выделяют гормон, который подавляет пищеварение у матери, пока они развиваются, потребляя очень большой запас желтка. Когда развитие лягушат заканчивалось, самка отрыгивала их наружу . Суринамская пипа выращивает своих детёнышей в порах на спине, где они остаются до метаморфоза . Сумчатая квакша после спаривания откладывает яйца, а самец помогает ей закинуть их на спину, где находится выводковая сумка. Через некоторое время из щели появляется 20—200 молодых лягушат. Ринодерма Дарвина — крохотная лягушка размером 3 см; самки откладывают яйца, в то же время самцы проглатывают их в резонатор под языком. Головастики растут несколько месяцев в резонаторах и вылазят через рот уже молодыми лягушатами . Поведение зернистой ядовитой лягушки Oophaga granulifera типично для целого ряда древесных лягушек семейства ядовитых древолазов Dendrobatidae. Её яйца откладываются на лесной подстилке, и когда они вылупляются, головастики переносятся один за другим на спине взрослой особи в подходящую заполненную водой щель, такую как свёрнутый лист или середина складок веток бромелии . Самка регулярно посещает «питомники» и откладывает неоплодотворённые яйца в воду, которые поедаются головастиками .
Большинство земноводных проводит жизнь во влажных местах, чередуя пребывание на суше и в воде, однако есть некоторые чисто водные виды, а также виды, проводящие жизнь исключительно на деревьях. Недостаточная приспособленность земноводных к обитанию в наземной среде обусловливает резкие изменения их образа жизни в связи с сезонными изменениями условий существования. Земноводные способны впадать в длительную спячку при неблагоприятных условиях (холода, засуха и т. п. ). У некоторых видов активность может меняться с ночной на дневную при понижении температуры ночью. Земноводные активны только в тёплых условиях. При температуре +7 — +8 °C большинство видов впадают в оцепенение, а при −2 °C — погибают. Но некоторые земноводные способны переносить длительное замораживание, пересыхание, а также регенерировать значительные утраченные части тела .
На суше земноводные ограничены влажными местами обитания из-за необходимости поддерживать кожу влажной . Некоторые земноводные, например жаба-ага Bufo marinus и крабоядная лягушка, Fejervarya cancrivor могут жить в солёной воде мангровых болот или, как саламандра Андерсона, в солоноватых и солёных озёрах . Однако жизнь большинства амфибий связана с пресной водой. Поэтому они отсутствуют на большинстве океанических островов, где условия для них в принципе благоприятны, но добраться до которых они самостоятельно не могут .


За некоторыми исключениями, взрослые современные земноводные во взрослой стадии — хищники, питающиеся практически всем, что движется, что они могут проглотить. Рацион в основном состоит из мелкой добычи, которая не двигается слишком быстро, такой как жуки, гусеницы, дождевые черви и пауки. Питаются амфибии мелкими животными (в основном насекомыми и беспозвоночными ), склонны к каннибализму . Растительноядных животных среди земноводных практически нет из-за крайне вялого обмена веществ . В рацион крупных водных видов может входить молодь рыб , а наиболее крупные могут охотиться на птенцов водоплавающих птиц и попавших в воду мелких грызунов . Сирены часто заглатывают водные растения вместе с беспозвоночными, которыми они питаются, а бразильская древесная лягушка ( ) ест большое количество фруктов . Мексиканская роющая жаба ( Rhinophrynus dorsalis ) имеет длинный язык для лова муравьёв и термитов. Она выбрасывает его кончиком вперёд, в то время как другие лягушки сначала выбрасывают заднюю часть языка. Крупные бесхвостые земноводные могут выбрасывать язык на 10 см в длину .

Земноводные являются первичными хищниками, важной составляющей
пищевой цепи
. Будучи
холоднокровными
животными, они эффективно используют потребляемую пищу, затрачивая лишь небольшую часть энергии на
метаболические
процессы и конвертируя остальное в
биомассу
. Они служат пищей вторичным хищникам, а сами питаются наземными членистоногими, в основном травоядными. Таким образом, поедая потребителей растений, амфибии увеличивают прирост биомассы растений, что способствует балансу
экосистемы
.
Пищу в основном выбирают на глаз, даже в условиях тусклого освещения. Движение жертвы вызывает атаку. Жабы, саламандры и червяги могут ориентироваться и на запах, чтобы обнаружить добычу. Эта реакция в основном вторична, потому что было замечено, что саламандры остаются неподвижными рядом с пахнущей добычей, а питаются только в том случае, если она движется. Пещерные амфибии обычно охотятся по запаху. Некоторые саламандры, кажется, научились распознавать неподвижную добычу, когда она не имеет запаха, даже в полной темноте .

Земноводные обычно глотают пищу целиком, но могут сначала слегка её пережёвывать. Они, как правило, имеют маленькие зубки на нёбе, это особенность, уникальная для амфибий. Их основание и коронка состоят из дентина, разделённого некальцинированным слоем, и они сменяются через определённые промежутки времени. Саламандры, червяги и некоторые лягушки имеют один или два ряда зубов в обеих челюстях, но у большинства лягушек отсутствуют зубы в нижней челюсти, а у жаб зубы вообще отсутствуют .
Тигровая саламандра, или тигровая амбистома ( Ambystoma tigrinum ) прячется под укрытием, устраивая засаду на неосторожных беспозвоночных. Другие амфибии, напротив, активно ищут добычу, в то время как аргентинская рогатая лягушка ( Ceratophrys ornata ) подманивает добычу ближе, поднимая задние лапы над спиной и вибрируя жёлтыми пальцами. Червяги ловят свою добычу, хватая её слегка направленными назад зубами. Борьба добычи и дальнейшие движения челюстей загоняют её внутрь, и червяк обычно отступает в свою нору. Пищу проглатывают целиком .
Личинки лягушек питаются желтком яйца. Когда он заканчивается, некоторые переходят на кормление бактериями, водорослевыми корками, детритом и стружкой с подводных растений. Вода втягивается через рот, который обычно находится в нижней части головы, и проходит через жаберные пищевые ловушки между ртом и жабрами, где мелкие частицы задерживаются в слизи и отфильтровываются. У других есть специализированный ротовой аппарат, состоящий из рогового клюва, окаймлённого несколькими рядами губных зубов. Они соскребают и кусают пищу многих видов, а также взбалтывают донный осадок, отфильтровывая более крупные частицы с помощью сосочков вокруг рта. У некоторых, таких как лопатоногая жаба, сильные кусачие челюсти, они плотоядны или даже каннибалы .

Характер питания личинок хвостатых земноводных практически аналогичен питанию взрослых животных. Личинки бесхвостых имеют кардинальное отличие, питаясь растительной пищей и детритом , переходя к хищничеству лишь к концу личиночной стадии .
Для размножения большинству земноводных требуется пресная вода, хотя некоторые откладывают яйца на суше. Некоторые могут обитать в солоноватой воде (например, крабоядная лягушка ), но морских амфибий нет. Однако несколько сотен видов лягушек, обычно обитателей тёплых областей, (например, Eleutherodactylus , Platymantis , живородящие жабы Nectophrynoides и многие другие тропические лягушки) не нуждаются в воде для размножения в дикой природе. Они размножаются живорождением, экологической и эволюционной адаптации, которая позволила им стать полностью независимыми от стоячей воды. Почти все эти лягушки живут во влажных тропических лесах, и из их икринок вылупляются уже взрослые лягушата, проходя через стадию головастика внутри яйца. Репродуктивный успех многих амфибий зависит не только от количества осадков, но и от времени года .
_mating_..._(35735771783).jpg)

В тропиках многие земноводные размножаются непрерывно или в любое время года. В регионах с умеренным климатом размножение в основном сезонное, обычно весной, и вызвано увеличением продолжительности дня, повышением температуры или дождём. Эксперименты показали важность температуры при размножении амфибий .
У червяг оплодотворение внутреннее, самец выдавливает сперму в клоаку самки. Парные мюллеровы железы внутри мужской клоаки выделяют жидкость, напоминающую жидкость, вырабатываемую предстательной железой млекопитающих, которая может транспортировать и питать сперму. Оплодотворение, вероятно, происходит в яйцеводе .
Большинство саламандр также размножаются внутренним оплодотворением. Самец проталкивает сперматозоиды в клоаку самки . В большинстве случаев самец откладывает сперматофор , небольшую капсулу спермы поверх желеобразного конуса, на субстрат либо на суше, либо в воде. Самка берёт пакет со спермой, захватывая его губами клоаки и проталкивая в отверстие. Сперматозоиды перемещаются в сперматеку на клоаку, где остаются до овуляции , которая может наступить через много месяцев. Ритуалы ухаживания и методы передачи сперматофора различаются у разных видов. У некоторых сперматофор может быть помещён непосредственно в клоаку самки, в то время как у других самка может направляться к сперматофору или удерживаться объятиями, называемыми амплексусом. Некоторые примитивные саламандры из семейств Sirenidae , Hynobiidae и Cryptobranchidae размножаются внешним оплодотворением так же, как и лягушки: самка откладывает яйца в воду, а самец выделяет сперму на яйцеклетку .
За некоторыми исключениями, лягушки размножаются внешним оплодотворением. Как правило, самцы обычно прибывают первыми к месту размножения, призывают кваканьем самок. Другие самцы спокойно остаются поблизости, ожидая возможности отобрать самку. Самки выбирают партнёра и откладываются яйца. Самец крепко обхватывает самку передними конечностями либо за передними, либо перед задними лапами, или вокруг шеи. Они остаются в амплексусе , их клоаки расположены близко друг к другу. Самка откладывает яйца, а самец покрывает их спермой. Шероховатые подушечки на лапках самца помогают удерживать хватку. Часто самец собирает и удерживает яичную массу, образуя задними лапами подобие корзины. Хвостатая лягушка размножается внутренним оплодотворением. «Хвост» есть только у самца и используется для оплодотворения самки. Эта лягушка живёт в ручьях с быстрым течением, и внутреннее оплодотворение сохраняет сперму, не унося её водой .

Большинство хвостатых амфибий ползают, волоча своё тело по земле. В воде они плавают, шевеля хвостом и телом, чем похожи на крокодила. Безногие земноводные передвигаются, как змеи или черви, волоча тело по земле, двигая им. Плоский и широкий череп сочленяется с позвоночником при помощи 2 мыщелков , образованных затылочными костями. У большинства земноводных четыре пальца на передней лапе и пять на задней, но ни на одной из них нет когтей (только у некоторых саламандр и волосатой лягушки ). У некоторых саламандр меньше пальцев. Безногие амфибии передвигаются на манер дождевых червей с перемещающимися вдоль тела зонами мышечных сокращений. На поверхности земли или в воде они передвигаются, покачивая телом из стороны в сторону . Лапы имеют приспособления к образу жизни: перепонки между пальцами для плавания, широкие клейкие подушечки на пальцах для лазания и ороговевшие бугорки на задних лапах для рытья (лягушки обычно копают землю назад). У большинства саламандр конечности короткие. Передвижение по суше осуществляется ходьбой, а хвост часто качается из стороны в сторону или используется в качестве опоры, особенно при лазании. При их обычной походке выдвигается вперёд только одна нога, как это было принято у их предков, лопастных рыб . Некоторые саламандры (многие виды рода лазающие саламандры ) лазают по деревьям и имеют длинные конечности, большие пальцы ног и цепкие хвосты . У водных саламандр и головастиков лягушек хвост имеет спинной и брюшной плавники и перемещается из стороны в сторону как средство движения. У взрослых лягушек нет хвостов, а у червяг они есть, однако они очень короткие . У бесхвостых же земноводных способов передвижения существенно больше . В зависимости от среды обитания они прыгают, бегают , ходят шагом , плавают , роют норы , лазают по деревьям или планируют . Бесхвостые считаются лучшими прыгунами из всех позвоночных (по длине прыжка относительно размеров тела) . Бесхвостые из семейств Bufonidae , Rhinophrynidae и Microhylidae имеют короткие задние конечности и передвигаются шагом, а не прыжками . Быстрое передвижение у представителей этих семейств обеспечивают либо ускоренные движения конечностей (быстрая ходьба), либо быстрые короткие прыжки . Бесхвостые, обитающие в воде, адаптированы к плаванию; для строения их тела характерны сильные задние конечности и наличие плавательных межпальцевых перепонок. Перепонки увеличивают площадь поверхности ступни (наподобие ласт) и способствуют быстрому передвижению лягушек в воде. Головастики, как правило, имеют большие хвостовые плавники, придающие ускорение, направленное вперёд, когда хвост движется из стороны в сторону. В воде бесхвостые наиболее беспомощны во время метаморфоза, когда хвост уже атрофирован, а ноги ещё не полностью функциональны . Некоторые бесхвостые адаптировались к жизни под землёй и, как следствие, к рытью нор. У этих видов бесхвостых, как правило, округлое туловище, короткие конечности, маленькая голова с выпученными глазами, а задние ноги адаптированы к рытью .
На протяжении своей эволюционной истории несколько не связанных между собой видов бесхвостых адаптировались к планирующему полёту .

Крики червяг и саламандр ограничиваются случайным тихим писком, ворчанием или шипением и мало изучены. Щёлкающий звук, который иногда издают червяги, может быть средством ориентации, как у летучих мышей, или формой общения. Большинство саламандр считаются безголосыми, но гигантская калифорнийская саламандра ( Dicamptodon ensatus ) имеет голосовые связки и может издавать дребезжащий или лающий звук. Некоторые виды саламандр издают тихий писк или визг при нападении .
Лягушки гораздо громче, особенно в период размножения, когда они используют свой голос для привлечения партнёров. Присутствие определённого вида в какой-либо местности легче определить по его характерному крику, чем по мимолётному взгляду самого животного. У большинства видов звук производится путём выброса воздуха из лёгких через голосовые связки в воздушный мешок или мешочки в горле или в углу рта. Он может раздуваться, как воздушный шар, и действовать как резонатор, помогая передавать звук в атмосферу или воду, когда животное находится под водой. Основная вокализация — это громкий, привлекающий самок крик самца. Этот призыв изменяется на более тихий к ухаживанию при приближении самки или если приближается другой самец-конкурент. Громкие звуки, в свою очередь, весьма опасны, так как могут привлечь хищников и требуют больших затрат энергии . Обычно ночная кубинская древесная лягушка ( Osteopilus septentrionalis ) издаёт громкий сигнал, когда идёт дождь в светлое время суток .

.jpg)
Мало что известно о территориальном поведении червяг, но некоторые лягушки и саламандры защищают свои участки территории. Обычно это места кормления, размножения или укрытия. Обычно такое поведение проявляют самцы, хотя у некоторых видов также участвуют самки и даже молодые особи. У некоторых из них есть особые приспособления, такие как увеличенные зубы для укусов или шипы на груди, лапах или больших пальцах. У саламандр защита территории включает в себя принятие агрессивной позы и, при необходимости, нападение на конкурента. Это может включать в себя хватку, погоню, а иногда и укусы, что иногда приводит к потере хвоста. Поведение красноспинных саламандр ( Plethodon cinereus ) хорошо изучено. 91 % отмеченных особей, которые позже были повторно пойманы, находились в пределах метра (ярда) от их первоначального дневного укрытия под бревном или камнем . Саламандры оставляли запаховые следы вокруг своих территорий, которые в среднем составляли от 0,16 до 0,33 квадратных метра (от 1,7 до 3,6 квадратных футов), и иногда на них жили пара самцов и самок. Саламандры поднимают тело, поддерживая себя с помощью хвоста и смотрят на противника. Обычно производился резкий выпад либо в область хвоста, либо в носогубные бороздки .
У лягушек территориальное поведение самцов часто наблюдается в местах размножения; призыв — это одновременно объявление о праве собственности на часть этого ресурса и привлекающий призыв к потенциальным самкам. В целом, более глубокий голос показывает более тяжёлую и сильную особь, и этого может быть достаточно, чтобы предотвратить вторжение более мелких самцов. Самцы обычно терпимо относятся к самцам-владельцам соседних территорий, в то же время энергично нападая на неизвестных особей. Если конкуренты не испугались предупреждающего крика, могут иметь место резкие выпады и толчки в грудь. Методы борьбы включают в себя толчки и пихания, сдувание голосового мешка противника, захват его за голову, прыжки на спину, укусы, преследование, брызги и погружение его под воду .


У земноводных мягкие тела с тонкой кожей, у них нет когтей, защитной брони или шипов. Поэтому, чтобы уберечься от хищников, многие из амфибий развили различные защитные механизмы. Первая линия защиты у саламандр и лягушек — это выделяемый ими слизистый секрет, который сохраняет кожу влажной и делает их скользкими и трудными для захвата. Секреция часто бывает липкой, неприятной или токсичной. Было замечено, что у змей происходит воспаление во рту, когда они пытаются проглотить африканских когтистых лягушек ( Xenopus laevis ), что даёт лягушкам возможность убежать . Червяги мало изучены в этом отношении, но кайенская водная червяга ( Typhlonectes compressicauda ) производит токсичную слизь, убившую хищную рыбу в эксперименте по кормлению в Бразилии. У некоторых саламандр кожа ядовита . Желтобрюхий тритон из Северной Америки и другие представители этого рода содержат нейротоксин тетродотоксин , почти идентичный тому, который содержится в рыбе-фугу . Обращение с тритонами не причиняет вреда, но проглатывание даже самых незначительных количеств кожи смертельно. В экспериментах было обнаружено, что рыбы, лягушки, рептилии, птицы и млекопитающие восприимчивы к яду тритонов . Единственными хищниками, обладающими некоторой толерантностью к яду, являются определённые популяции обыкновенных подвязочных змей ( Thamnophis sirtalis ). В местах, где сосуществуют и змеи, и саламандры, змеи выработали иммунитет благодаря генетическим изменениям и потому питаются амфибиями. Коэволюция происходит с тритоном, увеличивающим свои токсические способности с той же скоростью, с которой змея развивает свой иммунитет . Некоторые лягушки и жабы ядовиты, основные ядовитые железы находятся сбоку на шее и под бородавками на спине. Эти области представляются нападающему животному, и их выделения могут быть неприятными на вкус или вызывать различные физические или неврологические симптомы. Всего было выделено более 200 токсинов из ограниченного числа исследованных видов амфибий


Самые ядовитые позвоночные животные на Земле принадлежат к отряду бесхвостых земноводных — это лягушки- древолазы . Яд , который выделяют кожные железы земноводных, содержит убивающие бактерии вещества ( бактерициды ). У большинства земноводных России яд совершенно безвреден для человека. Однако многие тропические лягушки не так безопасны .
Абсолютным «чемпионом» по ядовитости среди всех позвоночных, включая змей, следует признать жительницу тропических лесов Колумбии — крохотного, размером всего 2—3 см, ужасного листолаза (местные жители называют его «кокои» ). Её кожная слизь содержит батрахотоксин . Из кожи кокои индейцы приготовляют яд для стрел. Одной лягушки достаточно, чтобы отравить 50 стрел. Два миллиграмма очищенного яда достаточно, чтобы убить человека .

Ядовитые виды часто используют яркую окраску, чтобы предупредить потенциальных хищников о своей токсичности. Эти предупреждающие цвета, как правило, красный или жёлтый в сочетании с чёрным, например, огненная саламандра. Как только хищник попробовал сьесть животное, он, вероятно, запомнит окраску. У некоторых видов, таких как огнебрюхая жаба ( Bombina spp. ), предупреждающая окраска находится на животе, и эти животные принимают оборонительную позу при нападении, демонстрируя хищнику свои яркие цвета. Лягушка Allobates zaparo не ядовита, а имитирование других токсичных видов в его местности — стратегия, которая может обмануть хищников ( мимикрия ) . В случае присутствия обоих видов вместе, Allobates zaparo копирует менее токсичную .
Многие амфибии ведут ночной образ жизни и днём прячутся, тем самым избегая дневных хищников, охотящихся на виду. Другие амфибии используют камуфляж, чтобы не быть обнаруженными. Они окрашены в различные цвета, такие как пятнистые, коричневые, серые и оливковые, которые сливаются с местностью. Некоторые саламандры принимают оборонительные позы, когда сталкиваются с потенциальным хищником, таким как североамериканская северная короткохвостая землеройка ( Blarina brevicauda ). Их тела корчатся, они поднимают и хлещут хвосты, из-за чего хищнику трудно избежать контакта с их гранулярными железами, вырабатывающими яд. Некоторые саламандры при нападении на них отбрасывают свой хвост, жертвуя этой частью тела, чтобы убежать . Хвост может иметь сужение у основания, чтобы его можно было легко отсоединить. Хвост регенерируется позже, но затраты энергии на его замену у животного значительны. Некоторые лягушки и жабы надуваются, чтобы казаться большими и свирепыми для змей, а некоторые чесночницы кричат и прыгают на нападавшего. Гигантские саламандры рода Andrias , а также лягушки из родов Ceratophrine и Pyxicephalus обладают острыми зубами и способны прокусить кожу человека или млекопитающего при оборонительном укусе . Некоторые виды совмещают маскировочную стратегию с апосематизмом. Так, краснобрюхая жерлянка имеет нейтральный окрас верхней части тела и яркие красные или жёлтые пятна на брюхе. В случае опасности жерлянка принимает позу, демонстрирующую яркие пятна, и отпугивает хищника .
В связи с тем, что органы тела снабжаются смешанной кровью, у амфибий низкий уровень обмена веществ , и поэтому они холоднокровные животные . Земноводные — холоднокровные, или пойкилотермные животные , температура тела которых не поддерживается за счёт внутренних физиологических процессов, а зависит от температуры внешней среды. Их скорость метаболизма низкая, и в результате их потребности в пище и энергии ограничены .

Земноводные встречаются на всех континентах, кроме Антарктиды, и в подавляющем большинстве наземных экосистем Земли. В некоторых типах экосистем (например, в смешанных лесах умеренной зоны Неарктики ) они являются одними из самых многочисленных (относительно других животных и по биомассе) позвоночных, уступая лишь грызунам. Как хищники они оказывают значительное влияние на пищевые цепи и поток энергии в экосистемах: на Ближнем Востоке увеличение вылова амфибий для пищевых целей привело к сокращению популяции лягушек и значительному увеличению численности количество комаров, численность которых ранее ограничивалась охотящимися на них амфибиями. Многие хищники, более сильные, полагаются на земноводных как на основной источник пищи. Стройная подвязочная змея питается почти исключительно двумя видами лягушек - иосемская жаба и . Сокращение их численности угрожает популяции этой змеи и других видов (включая хищных птиц). В мелких и периодических водоёмах головастики амфибий являются одними из основных детритофагов и травоядных, ими питаются различные хищные беспозвоночные (личинки и взрослые формы водяных скорпионов ), жуков-плавунцов , рыбы, птицы). Было подсчитано, что только в странах СНГ 91-109 видов птиц охотятся на бесхвостых .
.jpg)
Одним из самых опасных заболеваний земноводных является хитридиомикоз. Хитридиомикоз считается самой агрессивной и смертоносной инфекцией из когда-либо поражавших позвоночных, поскольку к нему восприимчиво большое количество видов и огромен урон, наносимый этим заболеванием. Её считают одним из факторов современного шестого массового вымирания . Одной из основных причин распространения этой инфекции по всему миру является международная торговля амфибиями, как декоративными видами, так и употребляемыми в пищу. Хитридиомикоз включён в список инфекций, контролируемых Всемирной организацией по охране здоровья животных .
.jpg)
.jpg)
У поражённых хитридиомикозом земноводных развиваются следующие физиологические патологии: эпидермальная дисфункция вследствие гиперкератоза эпидермиса (утолщение кожи приводит к нарушению обмена веществ с внешней средой, который у амфибий в значительной степени происходит через кожу, животные не могут получать необходимые питательные вещества, выделять токсины и нормально дышать), снижается уровень натрия и калия в крови и клеточной плазме, развивается дисбаланс электролитов , нарушается электролитический транспорт через эпидермис. Размножение Batrachochytrium dendrobatidis в тканях кожи хозяина (часто оно не приводит к воспалительной реакции со стороны организма хозяина) приводит к нарушениям её нормального функционирования — у большинства видов амфибий при достижении количества зооспор грибка до 10 000 штук заболевшие животные не могут нормально дышать, нарушается увлажнение, осморегуляция и терморегуляция . В крови заболевших земноводных снижается уровень определённых электролитов , таких как натрий , магний и калий . Негативно влияют на здоровье хозяина также и продукты жизнедеятельности грибков, действующие как токсины , снижающие общий иммунитет животного, влияющие на пролиферацию лимфоцитов и вызывающие апоптоз клеток. Гибель больного животного наступает через несколько дней после заражения .
Для бесхвостых земноводных также опасен Ribeiroia : род паразитических плоских червей из класса трематод , отращивающий захваченному земноводному дополнительную пару (или даже больше) задних лап, что делает заражённую лягушку лёгкой добычей для птиц, которые должны стать конечным хозяином для Ribeiroia .

Самки комаров рода Corethrella питаются кровью бесхвостых амфибий. Жертву они находят, ориентируясь по звуковым брачным сигналам лягушек. Самцы лягушек издают сложные и простые позывные сигналы. Сложные сигналы предпочитаются, как самками комаров, так и самками лягушек. Если самцы издают простые звуки, они меньше подвергаются нападению кровососов, но и вероятность спаривания таких самцов уменьшается. Комары служат переносчиками трипаносомозов лягушек .
Опасны также для хвостатых и бесхвостых земноводных лягушкоедки, или люцилии . Лягушкоедка — это зелёная падальная муха, паразитирующая на лягушках и жабах . В период размножения летом мухи откладывают свои белые продолговатые яйца на спину или голову жаб и других амфибий. Хозяевами паразита являются: обыкновенная , камышовая , зелёная жабы , травяная лягушка , жаба-повитуха , обыкновенная чесночница , обыкновенная квакша , остромордая лягушка , съедобная лягушка и даже огненная саламандра . Вылупившиеся личинки проникают через носовые отверстия в голову хозяина, где въедаются в мягкие ткани ещё живущего животного. Через несколько дней передняя часть морды и головы может полностью деформироваться и исказиться. Когда личинки добираются до мозга или других жизненно важных органов, функции организма нарушаются и амфибия погибает. В своей последней фазе личинки почти полностью съедают труп животного, включая кожу и соединительные ткани, при определённых обстоятельствах — даже хрящи и кости .
Специализированными хищниками на бесхвостых земноводных являются жуки и их личинки рода в составе семейства жужелиц .
Земноводные имеют способности привыкания, ассоциативного обучения как с помощью классического, так и инструментального обучения, а также способностей к различению количества .
В эксперименте, когда саламандрам предложили живых плодовых мушек ( ), они выбрали больше насекомых из 1 и 2 и из 2 и 3 особей. Лягушки могут различать небольшие количества (1 против 2, 2 против 3, но не 3 против 4) и большие количества (3 против 6, 4 против 8, но не 4 против 6) добычи .

В экстремальных погодных условиях некоторые виды бесхвостых погружаются в состояние анабиоза и могут не проявлять никакой активности в течение нескольких месяцев. В холодных областях бесхвостые зимой впадают в спячку . Некоторые наземные бесхвостые (например, американская жаба ) роют себе на зиму нору и спят в ней. Другие виды, менее приспособленные к рытью, прячутся в расщелинах или закапываются в сухие листья. Водные виды (например, лягушка-бык), как правило, залегают на дне водоёма, частично погружаясь в ил , но сохраняя, тем не менее, доступ к кислороду, растворённому в воде. Их метаболизм замедляется, и они выживают за счёт потребления внутренних энергетических резервов. Многие бесхвостые могут выжить после замерзания. Несмотря на то, что под их кожей и в полостях тела образуются кристаллы льда, жизненно важные органы защищены от замерзания вследствие высокой концентрации глюкозы в тканях. С виду безжизненная, замёрзшая лягушка может начать дышать и возобновить работу сердца, если её отогреть .

С другой стороны, роющая лопатница во время жаркого сухого сезона в Австралии впадает в состояние эстивации (летней спячки), обходясь без пищи и воды 9—10 месяцев в году. Эта лягушка зарывается в землю и сворачивается в защитном коконе, который образует её сброшенная кожа. Исследования показали, что во время эстивации метаболизм лягушки меняется так, что КПД митохондрий увеличивается, позволяя более эффективно использовать ограниченное количество энергетических ресурсов, доступных лягушке в состоянии спячки. Пытаясь ответить на вопрос, почему этот механизм не имеет широкого распространения в царстве животных, исследователи пришли к выводу, что он полезен только холоднокровным животным, находящимся в состоянии анабиоза в течение длительного периода, чьи энергетические затраты крайне низки, поскольку у них нет необходимости постоянно генерировать тепло . Другое исследование показало, что для удовлетворения минимальных энергетических требований у лягушки атрофируется большинство мышц (за исключением мышц задних конечностей) .
Сибирский углозуб также может переживать анабиоз: его метаболизм замедляется, и он выживает за счёт потребления внутренних энергетических резервов, а его жизненно важные органы защищены от замерзания вследствие высокой концентрации глюкозы в тканях .
Роющая лягушка может проводить от 8 до 10 месяцев в году в анабиозе, экономя воду и пережидая условия засухи и экстремальных температур. Она зарывается глубоко в землю, где покрывает себя слизью, выставляя наружу только ноздри и входит в состояние низкой метаболической активности и задержки воды. Как только период засухи заканчивается и идут дожди, бесхвостые земноводные выходят из анабиоза и вылезают на сушу, начиная поедать всё живое. Некоторые племена в Африке откапывают этих лягушек и пьют из них воду .
.jpg)
Благодаря живучести земноводные часто используются в качестве лабораторных животных .
Люди употребляют амфибий в пищу, выращивают их как домашних животных и т. п. Они естественным образом контролируют количество животных, считающихся вредителями (насекомые, улитки) , а некоторые виды амфибий массово использовались для борьбы с вредителями сельскохозяйственных культур (насекомыми, грызунами) . Для борьбы с вредителями посевов сахарного тростника на Карибские острова и в Австралию была завезена гигантская жаба яга ( Rhinella marina). В Карибском бассейне эта кампания имела умеренный успех, но в других частях света интродукция аги вызвала значительное снижение биоразнообразия — эта прожорливая амфибия питалась не только вредителями, но и другими видами и стала досаждать и уничтожать их . В связи с чувствительностью к изменениям условий обитания и наличием токсических веществ (в том числе из-за голой, проницаемой кожи, сложной, восприимчивой к нарушениям жизненного цикла) их используют как виды-индикаторы, свидетельствующие о состоянии экосистемы, а некоторые лягушки съедобны — их едят, в том числе, во Франции и на Дальнем Востоке. Лягушек и жаб также скармливают пушным зверям на фермах . Многие виды из семейства Dendrobatidae (преимущественно из рода Phyllobates ) используются индейцами Центральной и Южной Америки для отравления стрел, используемых для охоты. Присутствующие в них алкалоиды также являются предметом исследований в плане получения новых лекарственных средств (в том числе анестетиков) . Различные виды энтеогенов (главным образом буфотенин) получают из кожи и околоушных желёз земноводных во многих районах мира, в том числе среди племён Мезоамерики и Дальнего Востока . Когтистая лягушка, или же гладкая шпорцевая лягушка ( Xenopus laevis ) является важным модельным организмом, особенно в исследованиях эмбриогенеза и регуляции транскрипции (в том числе из-за её крупных эмбрионов, с которыми легко манипулировать, и прозрачных головастиков). Она оказала неоценимые услуги и в медицине: благодаря чувствительности к хорионическому гонадотропину особи этого вида послужили первым в истории медицины тестом на беременность (достаточно было ввести самке мочу женщины, присутствие хорионического гонадотропина стимулировало лягушка откладывает яйца через несколько часов). Аналогичным образом в качестве модельного организма используется родственный вид — . Жабы, равнинные и горные жабы ( Bombina bombina и B. variegata ) используются в качестве модельных организмов при изучении процессов межвидовой гибридизации и видообразования . Головастики многих видов амфибий, обитающих в тропиках и зонах умеренного климата, часто используются для изучения фенотипической изменчивости и индукции механизмов защиты от хищничества и механизмов формирования видового разнообразия . Кожа самых крупных лягушек идёт на украшения, сумки и другие изделия .
.jpg)
Земноводные отличаются среди других позвоночных разнообразием хромосом и геномов , а также их спецификой . Кариотипы были определены по крайней мере для 1193 (14,5 %) из ≈ 8500 известных видов: 963 бесхвостых, 209 хвостатых и 21 червяг. Как правило, кариотипы амфибий характеризуются 20—26 двуплечими хромосомами. Земноводные также имеют очень большие геномы по сравнению с другими видами позвоночных. Размеры генома колеблются от 0,95 до 11,5 пг у лягушек, от 13,89 до 120,56 пг у саламандр и от 2,94 до 11,78 пг у червяг .
Большие размеры генома препятствуют секвенированию всего генома земноводных, хотя недавно был опубликован ряд геномов. Проект по раскрытию генома лягушки Xenopus tropicalis объёмом 1,7 ГБ был первым для земноводных в 2010 году. По сравнению с некоторыми саламандрами геном лягушки крошечный. Например, геном мексиканского аксолотля оказался размером 32 Гб, что более чем в 10 раз больше генома человека (3 Гб) . Размер мейотических бивалентов у хвостатых амфибий может достигать 700 мкм, как у огненной саламандры ( Salamandra salamandra) и обыкновенного тритона Lissotriton vulgaris (ранее Triturus vulgaris ) .
В Proceedings of the National Academy of Sciences (PNAS) в 2021 году была опубликована статья, обобщающая результаты многолетнего исследования гибридизации земноводных. Авторы проанализировали зоны гибридизации 41 пары европейских амфибий и измерили проницаемость всего генома. Цель состояла в том, чтобы оценить, возникает ли репродуктивная изоляция как быстрый процесс, вызванный мутациями в нескольких ключевых генах, или скорее как медленный процесс, включающий множество случайных мутаций, разбросанных по всему геному .
Результаты показывают, что репродуктивная изоляция у амфибий изначально развивается за счет множественных генетических несовместимостей, распространяющихся по всему геному, а не только по нескольким ключевым генам. Когда линии молодые, они могут без проблем скрещиваться, иногда на больших географических расстояниях — более ста километров для подвида одного вида лягушек в Каталонии. Миллионы лет спустя несовместимости затрагивают больше генов, то есть постепенно разрушаясь, и обмен происходит только в нескольких километрах — как между жабами на юге Польши. Таким образом, эти открытия дают новое объяснение тому, почему виды земноводных гораздо древнее других позвоночных. Репродуктивная изоляция может развиваться медленнее у амфибий, потому что требуется время, чтобы накопить множество несовместимостей, необходимых для ограничения гибридизации .



Сокращения популяций земноводных задокументированы для большинства видов земноводных. Данные проведённых мониторингов многих видов по всему миру свидетельствуют о сокращениях или даже полном исчезновении ряда популяций. На основании этого возникла распространённая гипотеза о глобальном сокращении численности земноводных, которое отмечается с конца 1980-х годов в различных местах по всему миру, и считается одной из наиболее серьёзных угроз глобальному биоразнообразию . В 2006 году считалось, что 2035 видов земноводных находятся под угрозой исчезновения, и эта цифра, вероятно, занижена, поскольку она не включает 1427 видов, по которым не было достаточно данных для оценки их статуса . Считается, что это связано с рядом причин, включая разрушение и изменение среды обитания, загрязнение почвы и воды, глобальное потепление, загрязняющие вещества, нарушающие работу эндокринной системы земноводных, разрушение озоного слоя (показано, что ультрафиолетовое излучение особенно вредно для кожи, глаз и яиц земноводных) и такие заболевания, как хитридиомикоз . Антропогенные факторы рассматриваются главной причиной сокращений популяций земноводных в историческое время. Обезлесение приводит к сильному изменению и разрушению биотопов суши, пересыханию водоёмов, используемых земноводными для размножения. Наиболее уязвимыми являются лесные и горные виды. Второе место занимает осушение либо загрязнение водоёмов, главным образом пестицидами и синтетическими удобрениями, что вызывает уничтожение биотопов, где проходит развитие наиболее уязвимых стадий жизненного цикла (эмбрионов и личинок), а также приводит к фрагментации популяций. Наиболее опасным является промышленное загрязнение водоёмов, поскольку земноводные на личиночных и постметаморфозных стадиях могут накапливать в своих организмах тяжёлые металлы . Негативно на численности популяций сказывается и урбанизация. Усиленный либо массовый отлов (особенно для террариумистики) также может быть фактором сокращения популяций . Из прочих факторов важную роль играют интродуцированные хищники. Например, в ряде мест на Кавказе енот-полоскун стал массовым хищником, истребляющим местные виды земноводных. Большое влияние на численность популяций земноводных в Европе оказывает ротан , интродуцированный в начале XX века. Чернобрюхая дискоязычная лягушка считалась вымершей с 1943 года и была вновь обнаружена в 2011 году . Синантропные млекопитающие (собаки, кошки, лисы) и птицы (серые вороны, сороки) представляют угрозу земноводным, обитающим на территории населенных пунктов и их окрестностей . Некоторые причины сокращения численности амфибий до сих пор неизвестны и являются предметом постоянных дискуссий .
Группа специалистов по земноводным из МСОП возглавляет усилия по реализации глобальной стратегии сохранения земноводных . Ковчег амфибий — это организация, которая была создана для реализации рекомендаций по сохранению земноводных, и они работают с зоопарками и аквариумами по всему миру. Одним из таких проектов является проект по спасению и сохранению амфибий в Панаме. Была разработана программа «Панамский проект по спасению и сохранению амфибий» ( англ. The Panama Amphibian Rescue and Conservation Project ) для сохранения некоторых видов бесхвостых в восточной Панаме , гибнущих от этих заболеваний, включающая разработку полевых методов использования пробиотиков .
.jpg)

Земноводные играют важную роль в культуре, но, как отмечает исследователь культуры народов мира Мариан Млынарски, они, как и рептилии , не самые известные животные. Это способствовало формированию ложного представления об амфибиях: как правило, они считаются отталкивающими существами, вызывающими страх или отвращение. Это совершенно необоснованное мнение имело негативные последствия, в том числе и бездумное истребление этих животных .

С земноводными связано множество мифов и суеверий. Например считалось, что некоторые саламандры могут выживать в огне и даже жить в нём, как в среде обитания . Протеи , в свою очередь, в Словении XVIII века считались детьми драконов , живущими в центре земли . В прошлом древесным лягушкам приписывали способность предсказывать погоду, и они считались полезными в составлении прогнозов. По этой причине её содержали в специальных высоких террариумах. Их и сейчас иногда содержат в качестве домашних питомцев, а также популярную в разведении изумрудную австралийскую квакшу .
В библейской Книге Исход размножение жаб является вторым бедствием в Египте .
Ян Парандовски называет Лернейскую гидру , чудовище болот из древнегреческой мифологии, «отвратительной амфибией» . Слово «тритон», которым сегодня описывают тритонов, происходит из греческой мифологии. Это было имя одного морского бога, сына Посейдона, и оно означало целую расу морских существ , наполовину людей, наполовину чудовищ. В корейской мифологии есть правитель по имени Кымва, или «Золотая лягушка» . Названия мест и организаций также связаны с амфибиями .
Волосатая лягушка заняла первое место в списке самых странных животных в мире, составленном сайтом New Scientist . Благодаря своим причудливым «когтям» её сравнивают с героями комиксов и фильмов, например с Росомахой .
Иногда науку о земноводных называют батрахологией (от др.-греч. βάτραχος — лягушка ), однако официально признанной такой науки нет . В первую очередь это связано с тем, что узких специалистов исключительно по амфибиям практически нет. Признанная наука о амфибиях и рептилиях — это герпетология. Она, в свою очередь, является частью более широкой науки: зоологии, хотя редко земноводные расматриваются в её рамках. Исследования в этой области касаются таксономии , морфологии , этологии , экологии , физиологии и биохимии животных . Результаты исследований подлежат научному рецензированию и публикации в научных журналах. Профессионалы проводят полевые работы, исследования в университетах, обучают студентов, управляют музеями и зоопарками, разводят и продают земноводных и управляют дикой природой. Они объединяются в научные общества (например, (SSAR) и (SEH)) и публикуют статьи в научных журналах, посвящённых герпетологии (например, ). Одним из самых крупных герпетологических обществ является « ». Это крупнейшее международное герпетологическое общество, признанное во всем мире за самую разнообразную программу публикаций, встреч и других мероприятий .
{{
cite journal
}}
: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (
ссылка
)
| Земноводные | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Тип:
Подтип:
Инфратип:
Надкласс:
Класс:
Земноводные
|
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Amphibia Linnaeus , 1758 | |||||||||||||||||||||||||||||||||||||||||
| Современные отряды | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Современный ареал | |||||||||||||||||||||||||||||||||||||||||

|
|||||||||||||||||||||||||||||||||||||||||
|
Геохронология
появился 345 млн лет
◄
Наше время
◄
Мел-
палеогеновое вымирание
◄
Триасовое вымирание
◄
Массовое
пермское вымирание
◄
Девонское вымирание
◄
Ордовикско-силурийское вымирание
◄
Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||