Стволовые клетки
- 1 year ago
- 0
- 0

Дендри́тные кле́тки (ДК, англ. Dendritic cells, DC ) — специализированные на презентации антигенов лейкоциты , имеющие характерную отростчатую морфологию. Дендритные клетки относят к «профессиональным» антигенпрезентирующим клеткам . Дендритные клетки играют важнейшую роль в функционировании иммунной системы , поскольку они необходимы для активации T-клеточного ответа. Дендритные клетки экспрессируют молекулы главного комплекса гистосовместимости I ( ) и II ( ), на которых представляют фрагменты антигенов T-клеткам. Без участия антигенпрезентирующих клеток T-клетки не могут распознать антиген.
Почти все дендритные клетки имеют костномозговое происхождение, за исключением фолликулярных дендритных клеток , имеющих мезенхимальное происхождение. Дендритные клетки костномозгового происхождения могут относиться как к миелоидному , так и лимфоидному ряду. Дендритные клетки можно получить из фибробластов , а также из моноцитов периферической крови или стволовых клеток костного мозга . Существует несколько разновидностей дендритных клеток: миелоидные дендритные клетки, плазмоцитоидные дендритные клетки , фолликулярные дендритные клетки и клетки Лангерганса .
Дендритные клетки играют важную роль в развитии и терапии аутоиммунных заболеваний , рака и других болезней. Для ряда вирусов , например, ВИЧ , дендритные клетки выступают в роли мишени. Иногда дендритные клетки сами становятся причиной рака и претерпевают злокачественное перерождение.
Дендритные клетки впервые описал Пауль Лангерганс в 1868 году. Впоследствии найденную им разновидность дендритных клеток назвали клетками Лангерганса. Функциональную роль дендритных клеток удалось установить во второй половине XX века. В 1973 году Ральф Стайнман и ввели в употребление термин «дендритные клетки». В 2011 году за открытие дендритных клеток Стайнману была присуждена Нобелевская премия по физиологии или медицине .
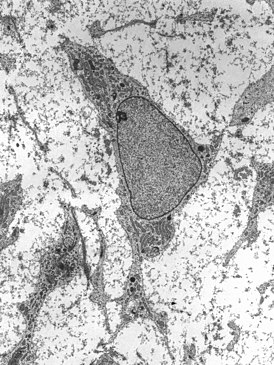
Дендритные клетки представляют собой подвижные крупные ( диаметром 15—20 мкм ) клетки овальной или многоугольной формы с эксцентрически расположенным ядром и длинными цитоплазматическими выростами , причём некоторые цитоплазматические выросты превышают 10 мкм. Цитоплазма дендритных клеток содержит небольшое количество внутриклеточным органелл , но достаточно большое количество митохондрий . Отростчатая морфология, наличие псевдоподий и ворсинок присущи всем зрелым дендритным клеткам независимо от их специализации. Именно наличию отростков дендритные клетки обязаны своим названием . К свойствам, универсальным для всех дендритных клеток, можно также отнести повышенный уровень экспрессии главного комплекса гистосовместимости I ( ) и II ( ) классов, а также молекул CD80 и CD86 . Кроме того, все дендритные клетки могут путём пиноцитоза или, в меньшей степени, фагоцитоза захватывать антиген для дальнейшего и презентации его T-клеткам, что приводит к активации последних .
В крови здоровых людей дендритные клетки имеют малый уровень экспрессии молекул CD83 , CD80, CCR7 , независимо от типа клетки .

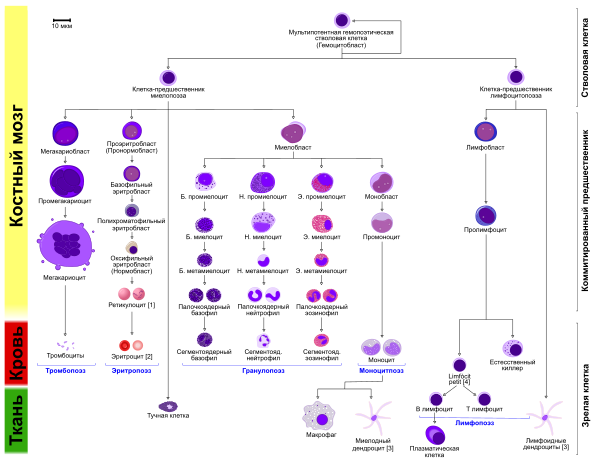
Начало дендритным клеткам дают кроветворные стволовые клетки , то есть дендритные клетки, за исключением фолликулярных дендритных клеток, имеющих мезенхимальное происхождение, происходят из костного мозга. Дендритные клетки образуются из клеток-предшественников как миелоидного, так и лимфоидного ряда, причём дифференцировка предшественников в дендритные клетки возможна даже на ранних стадиях развития. В то же время допускается существование особой клетки-предшественницы дендритных клеток .
Большинство дендритных клеток принадлежат к миелоидному ряду. Считается, что in vivo дендритные клетки могут образовываться как из гранулоцитарно-моноцитарных предшественников , так и из моноцитов . В культуре миелоидные дендритные клетки можно получить и из клеток костного мозга в присутствии GM-CSF и других цитокинов , а также из моноцитов в присутствии GM-CSF и IL-4 . Дендритные клетки лимфоидного ряда дифференцируются из общего лимфоидного предшественника, а также предшественников T- и B-клеток , в частности, на самой ранней стадии их развития в тимусе (DN1-клеток). Предшественников лимфоидных и миелоидных дендритных клеток объединяет экспрессия FLT3 , что отличает их от предшественников других иммунных клеток . Таким образом, дендритные клетки могут образовываться из 6—7 клеточных источников .
В периферической крови присутствуют незрелые дендритные клетки, которые составляют менее 0,5 % от общего числа лейкоцитов в кровотоке и выступают предшественниками дендритных клеток миелоидного и лимфоидного рядов, а также клеток Лангерганса. клеток-предшественников миелоидных клеток являются молекулы CD11c и MHC-II. Большинство незрелых дендритных клеток кровотока составляют так называемые плазмоцитоидные дендритные клетки, которые относятся к лимфоидному ряду. Незрелые дендритные клетки мигрируют из периферической крови в ткани благодаря тому, что на их поверхности располагаются рецепторы к практически всем β- хемокинам . Хотя дендритные клетки присутствуют во многих тканях и органах , их количество мало, поэтому и открыты они были сравнительно поздно. Как и макрофаги, дендритные клетки подразделяют на резидентные, которые присутствуют преимущественно в барьерных тканях (в коже и слизистых оболочках ), и воспалительные .
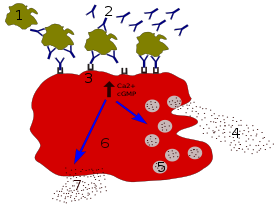
Основная функция дендритных клеток — презентация антигенов T-лимфоцитам, поэтому их относят к «профессиональным» антигенпрезентирующим клеткам. По эффективности презентации они на два порядка превосходят макрофаги благодаря повышенной экспрессии MHC, особенно MHC-II, а также молекул-костимуляторов. Презентация антигена дендритными клетками запускает иммунный ответ .
Дендритные клетки активно и непрерывно распознают патогены (точнее, , PAMP ) с помощью Toll-подобных рецепторов и поглощают соответствующие антигены по пути пиноцитоза и, в меньшей степени, фагоцитоза . Если чужеродных антигенов вокруг дендритной клетки нет, то она поглощает и презентирует T-клеткам материал собственных тканей организма, но без корецепторного стимула, и иммунный ответ против собственных тканей организма не развивается. До активации антигеном дендритные клетки остаются незрелыми и имеют низкую презентирующую способность. Созревание дендритной клетки происходит при распознавании патогена Toll-подобными рецепторами. Дендритные клетки миелоидного ряда, подобно макрофагам, несут рецепторы маннозы , липополисахарида , а также TLR2 и TLR4 , позволяющие распознавать грамотрицательные и грамположительные бактерии соответственно. Плазмоцитоидные дендритные клетки несут TLR7 и TLR9 , распознающие ДНК вирусов и бактерий .
После распознавания патогена в дендритных клетках увеличивается продукция цитокинов и хемокинов, а также повышается экспрессия адгезивных молекул , необходимые для представления антигенов и стимуляции Т-клеток . В большинстве случаев фрагменты процессированного антигена представляются дендритными клетками в комплексе с MHC-II. В отличие от других антигенпрезентирующих клеток дендритные клетки могут презентовать антиген цитотоксическим CD8 + T-лимфоцитам в комплексе с MHC-I. Этот процесс известен как . Одна зрелая дендритная клетка может активировать от 100 до 3 тысяч T-клеток .
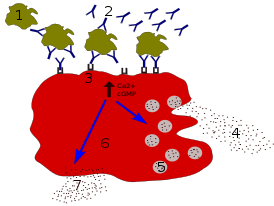
Под действием хемокинов , и CXCL12 зрелые дендритные клетки мигрируют из покровных тканей и по тканевой жидкости попадают в лимфатические сосуды , откуда перемещаются в региональные лимфоидные органы , например, в паракортикальную зону лимфатических узлов . В ходе миграции дендритная клетка практически утрачивает способность к эндоцитозу и проводит процессинг антигена с дальнейшим представлением его фрагментов в составе MHC. Попадание дендритных клеток в T-зависимую зону лимфатических узлов обеспечивается изменением экспрессии рецепторов хемокинов. Вместо рецепторов для хемокинов, которые продуцируются клетками барьерных тканей, дендритные клетки начинают экспрессировать рецепторы CCR7 и CXCR4 , которые распознают хемокины, выделяемые клетками стромы T-зависимых зон лимфатических узлов. В этих зонах происходит презентация антигенов дендритными клетками T-лимфоцитам и активация последних через корецепторы CD40 , CD80 и CD86. Контакт дендритной клетки и T-клетки через презентированный на MHC-II антиген получил название иммунологического синапса . Без иммунологического синапса дальнейшее развитие иммунного ответа невозможно . Именно большое количество MHC-II, а также CD80 и CD86 на поверхности зрелых дендритных клеток делает их антигенпрезентирующими . Активированные дендритные клетки продуцируют CD154 , который связывается с CD40 на поверхности дендритных клеток . Зрелые дендритные клетки, доставившие антиген в T-зависимую зону лимфоузла, становятся частью его стромы и называются интердигитальными дендритными клетками, так как между их отростками, напоминающими пальцы, располагаются T-лимфоциты .
Резидентные миелоидные дендритные клетки заселяют органы на стадии клеток-предшественников или незрелых дендритных клеток и завершают своё развитие уже в органе, который в дальнейшем не покидают. Многие резидентные дендритные клетки располагаются в тимусе, зародышевых центрах , селезёнки , печени и других органах .
Клетки эпителия миндалин и желудочно-кишечного тракта продуцируют хемокин , который привлекает незрелые дендритные клетки за счёт взаимодействия с CCR6 на поверхности незрелых дендритных клеток. Синтез этого хемокина активируется при воспалении. При созревании дендритные клетки перестают экспрессировать CCR6 и перестают реагировать на MIP-3α, но на их поверхности появляется другой хемокиновый рецептор — CCR7 . Созревающие дендритные клетки начинают реагировать на хемокины MIP-3β и , продуцируемый вторичными лимфоидными тканями, что привлекает дендритные клетки в лимфоузлы .
Некоторые факторы могут системно изменять и даже нарушать функционирование дендритных клеток. Так, дислипидемия изменяет функцию презентации антигена, а никотин повреждает их способность начинать пролиферацию Т-клеток и вырабатывать цитокины .
Зрелые дендритные клетки подразделяют на два типа: DC1 (миелоидные) и DC2 (лимфоидные). Основные свойства клеток этих типов представлены в таблице ниже :
| Характеристика | DC1 | DC2 |
|---|---|---|
| Происхождение | Миелоидные | Лимфоидные |
| Индуцирующие цитокины | , IFNγ | IL-10 |
| Фенотип | CD11c hi CD123 lo | CD11c - CD123 hi |
| Продуцируемые цитокины | IL-12 | IFNγ |
| Влияние на T-клетки | Активация | Индукция |
Незрелые лимфоидные дендритные клетки представлены плазмоцитоидными дендритными клетками. Особняком стоят эпидермальные дендритные клетки, известные как клетки Лангерганса, и фолликулярные дендритные клетки, имеющие мезенхимальное, а не костномозговое происхождение. В тимусе присутствуют в основном лимфоидные дендритные клетки, но есть и миелоидные. В селезёнке и брыжеечных лимфатических узлах преобладают в разных соотношениях популяции миелоидных лимфоидных клеток. В лимфатических узлах, связанных с кожей, помимо миелоидных дендритных клеток в больших количествах присутствуют клетки Лангерганса .
Согласно альтернативной классификации, дендритные клетки подразделяются на классические и плазмоцитоидные (см. ниже). Классические дендритные клетки стимулируют T-клеточный ответ. Именно классические дендритные клетки являются преобладающей группой дендритных клеток в лимфоидных тканях. Классические дендритные клетки дополнительно подразделяются на основные и кросс-презентирующие . Плазмоцитоидные дендритные клетки — это незрелые клетки, которые происходят от того же предшественника, что и классические дендритные клетки, и локализованы в основном в кровотоке, в меньшей степени — в лимфоидных органах. В отличие от классических дендритных клеток, плазмоцитоидные дендритные клетки практически неспособны к фагоцитозу и не поглощают антигены. Основной функцией плазмоцитоидных клеток является продукция интерферона .
Миелоидные дендритные клетки (МДК) — дендритные клетки-предшественницы гемопоэтического происхождения. МДК имеют схожую морфологию, а также эндоцитозную и ферментативную активность с макрофагами и моноцитами. Локализируются в эпидермисе, дерме , дыхательных путях , кишечнике , тимусе , селезёнке, печени и лимфоидной ткани. Они характеризуются низким уровнем экспрессии MHC, а также костимуляторных и адгезивных молекул. С помощью пиноцитоза и фагоцитоза МДК захватывают чужеродные антигены, после чего экспрессируют антигенную детерминанту в комплексе с молекулами MHC-II. Затем они мигрируют в местные лимфатические узлы, где стимулируют пролиферацию и дифференцировку антигенспецифических Т-лимфоцитов, провоцируя и стимулируя иммунный ответ. В ответ на стимуляцию индукторами созревания, МДК продуцируют преимущественно цитокины Т-Хелпера 1 типа, включая IL-6 , IL12RB1 , фактор некроза опухоли и интерферон γ. Маркерами МДК являются молекулы CD1c и CD141 .
Плазмоцитоидные дендритные клетки (ПДК) — незрелые клетки лимфоидного происхождения, морфологически напоминают плазматические клетки . ПДК — преобладающая разновидность незрелых дендритных клеток, циркулирующих в крови. ПДК меньше моноцитов (8—10 мкм), их ядро имеет менее выраженную выемку. Помимо кровотока, ПДК локализируются в Т-клеточных зонах лимфоидных тканей, миндалинах , тимусе, печени, лёгких и в коже. В присутствии IL-3 и бактериальных антигенов ПДК дифференцируются в зрелые лимфоидные дендритные клетки. На своей поверхности ПДК не несут молекул CD83 , CD11b и CD11c, которые характерны для миелоидных дендритных клеток, а также CD13 и CD14 , характерные для большинства миелоидных клеток. В ПДК экспрессируются гены , которые необходимы для перестройки генов антигенраспознающих рецепторов. На поверхности ПДК присутствуют CD45R0 и рецепторы для IL-3 , при этом MHC-II представлены в меньшей степени, чем на миелоидных дендритных клетках, причём молекулы MHC-II есть не только на поверхности, но и в цитоплазме. В геномах ПДК имеются признаки перестройки генов T-клеточных рецепторов , которые характерны для T-клеток. ПДК выделяют в больших количествах (α, β и ω), являясь основными поставщиками интерферона клеткам крови, поэтому ПДК также известны как клетки-продуценты интерферона. ПДК секретируют большие количества интерферонов преимущественно на первые сутки после стимуляции нуклеиновыми кислотами вирусного происхождения. Также они ответственны за производство цитокинов IL-4 и IL-10 , которые переключают дифференцировку наивных Т-хелперов в Т-хелперы 2 типа. В спектре Toll-подобных рецепторов, экспрессируемых ПДК, преобладают рецепторы, находящиеся на цитоплазматических гранулах и распознающие нуклеиновые кислоты. В частности, экспрессия TLR9, лигандами которого являются Cp G - олигонуклеотиды бактериальной ДНК , позволяет им обнаруживать патогены. Маркерами ПДК являются молекулы CD303 , CD304 , CD141, рецептор IL-3 CD123, а также иммуноглобулиноподобные рецепторы CD85k и .
ПДК попадают в лимфатические узлы тем же путём, что и T-клетки, а именно, через высокий эндотелий пост капиллярных венул . В то же время клетки-предшественницы миелоидных дендритных клеток проникают в лимфоузлы вместе с афферентным током лимфы . Как упоминалось выше, в течение первых суток после стимуляции вирусами или IL-3 ПДК усиленно экспрессируют интерфероны. В течение вторых суток они дифференцируются в зрелые дендритные клетки лимфоидного ряда. По мере созревания на поверхности ПДК повышается количество молекул MHC-II, появляются молекулы-костимуляторы CD80 и CD86. Во время созревания ПДК продолжают выделять интерфероны, но в меньшем количестве. При вирусной стимуляции созревающая ПДЦ способствует дифференцировке T-хелперов 1 типа (Th1-клеток), продуцирующих интерферон γ, а при стимуляции IL-3 — T-хелперов 2 типа (Th2-клеток), продуцирующих IL-4 .
Фолликулярные дендритные клетки (ФДК) — зрелые клетки, в отличие от остальных дендритных клеток, имеют мезенхимальное происхождение. Прочно соединяясь десмосомами отростков, образуют стабильную сеть, благодаря которой они не мигрируют в другие области. ФДК локализируются в клеточных зонах лимфатических узлов и селезёнке. Они представляют антиген В-лимфоцитам во вторичных лимфоидных органах. Антиген на поверхности ФДК пассивен, сохраняет исходную конформацию антигена с антителом . Эти клетки являются особым микроокружением, на котором происходит уникальный для В-лимфоцитов процесс соматической гипермутации вариабельных сегментов генов, кодирующих антитела, и отбор наиболее аффинных к антигену вариантов антител. Они не экспрессируют белки MHC-II, но несут много молекул Fcγ-рецепторов , и CR2 , что позволяет им связывать и представлять содержащийся в комплексах антиген В-лимфоцитам. При повторном поступлении антигена в организм, когда он связывается с ранее наработанными антителами, ФДК стимулируют B-клетки памяти .


Клетки Лангерганса — эпидермальные дендритные клетки миелоидного ряда. Своё название клетки Лангерганса получили в честь первооткрывателя Пауля Лангерганса , который описал их в конце XIX века как отростчатые клетки эпидермиса, однако их принадлежность к иммунным клеткам была установлена позднее. Клетки Лангерганса имеют ряд признаков, отличающих их от всех остальных дендритных клеток. В их цитоплазме присутствуют слоистые включения, известные как , а на поверхности присутствует рецептор лангерин ( CD208 ), относящийся к лектинам и присутствующий уже на циркулирующих в кровотоке клетках-предшественниках, и «неклассическая» молекула MHC — CD1a , которая предназначена для презентации липидных антигенов. Гранулы Бирбека представляют собой овально вытянутые структуры с центрально расположенным фрагментированным стержнем, происходящие из атипичных гранул, и напоминают по форме теннисные ракетки. Гранулы Бирбека, возможно, являются эндосомами , участвующими в захвате антигена . Атипичные гранулы клеток Лангерганса морфологически отличаются от лизосом наличием прозрачного ореола, разделяющего наружную мембрану гранулы и её центральную зону . Также в цитоплазме клеток Лангерганса присутствует трубчато-везикулярная система — соединение модифицированного и гипертрофированного комплекса Гольджи с гладким эндоплазматическим ретикулумом . Кроме упомянутых выше лангерина и CD1a, на поверхности клеток Лангерганса присутствуют такие молекулы, как Е-кадгерин , CD20 , CD11c , CD13 , CD14 и CD33 . Развитие клеток Лангерганса до конца не изучено. Считается, что они развиваются преимущественно из гемопоэтических клеток желточного мешка , а также, хотя и в меньшей степени, из костномозговых предшественников . Клетки Лангерганса локализируются в эпидермисе, сальных и потовых железах , волосяных фолликулах , эпителии полости рта, носа, пищевода , лёгких, влагалища и толстой кишки . Основная функция клеток Лангерганса — регуляция гомеостаза всего организма и реализация защитных реакций кожи . С помощью отростков они связаны с нервными окончаниями эпидермиса и с гемомикроциркуляторным руслом сосочкового слоя дермы . Благодарю этому на клетках Лангерганса замыкаются 3 регуляторные системы — иммунная, нервная и эндокринная . Уникальная особенность Клеток Лангерганса — сохранение антигена патогена в иммуногенной форме длительное время . После захвата антигена клетки Лангерганса перемещают антигенные пептиды в местные лимфатические узлы через центральные лимфатические сосуды и участвуют в дифференцировке наивных Т-лимфоцитов в Т-хелперы .
Дендритные клетки задействованы в иммунном ответе организма на многие вирусы и могут поражаться некоторыми из них. Так, клетки Лангерганса участвуют в быстром ВИЧ -инфицировании. Клетки экспрессируют рецепторные структуры для ВИЧ — CD4 и рецепторы хемокинов. После захвата антигена или инфицирования ВИЧ через 30 минут клетки Лангерганса мигрируют в лимфатические узлы, благодаря чему они способствуют быстрому распространению ВИЧ по организму . Дендритные клетки обеспечивают выработку интерферонов при заражении такими вирусами, как цитомегаловирус и вирус гриппа . Хантавирус при попадании в организм эффективно заражает дендритные клетки, при этом не провоцируя лизис или апоптоз . Хантавирус активирует незрелые клетки, вследствие чего повышается экспрессия MHC, а также костимулирующих и адгезивных молекул. Инфицированные хантавирусом клетки усиливают высвобождение , в частности, фактора некроза опухоли и α-интерферона. Таким образом, хантавирусы вызывают сильный иммунный ответ, являющийся частью патогенеза вирусной инфекции . Показано, что фолликулярные дендритные клетки могут способствовать размножению прионов при скрейпи .
Дендритные клетки вовлечены в патогенез ряда аутоиммунных заболеваний . У больных системной красной волчанкой активность дендритных клеток патологически повышена, вследствие чего повышается выделение интерферонов, стимулирующих дифференцировку клеток-предшественниц дендритных клеток. Зрелые дендритные клетки поглощают ДНК, содержание которой в крови при системной красной волчанке увеличивается, и начинается иммунный ответ: в организме образуются антитела против его собственной ДНК, которые и обуславливают заболевание . Показана роль дендритных клеток в развитии аллергии , а также , таких как болезнь Крона и язвенный колит .
Иногда дендритные клетки претерпевают злокачественное перерождение. Например, саркомы , происходящие из фолликулярных дендритных клеток, обычно затрагивают лимфоидные ткани, но также могут поражать печень, жёлчные протоки , поджелудочную железу , щитовидную железу и другие органы . Повышенное содержание дендритных клеток в опухоли коррелирует с лучшим прогнозом, таким образом, дендритные клетки могут оказывать влияние на развитие опухолей . Дендритные клетки активируют T-хелперы и цитотоксические T-клетки, которые могут подавлять развитие опухоли. Кроме того, дендритные клетки могут влиять на успех иммунотерапии рака , в частности, основанной на применении .
Способность дендритных клеток управлять иммунными процессами, а также успехи в разработке систем для получения клеток с заданными параметрами привели к их использованию в иммунотерапии рака и некоторых аутоиммунных заболеваний , а также атеросклероза . Иммунотерапия рака, направленная на дендритные клетки, была успешно протестирована на пациентах с гепатоцеллюлярной карциномой , карциномой почек , меланомой и лимфомами . Использование активации дендритных клеток в противораковой иммунотерапии основано на их способности активировать иммунный ответ против опухоли . Чтобы активировать дендритные клетки, используют цитокины, сшитые с опухолевыми антигенами . Как правило, в опухолях дендритных клеток немного, однако повышенная плотность популяций дендритных клеток считается маркером благоприятного прогноза . Дендритные клетки, входящие в состав опухолей, могут активировать T-хелперы и цитотоксические T-клетки, которые, в свою очередь, могут подавлять рост опухоли .
Благодаря способности модулировать силу иммунного ответа дендритные клетки могут быть использованы в терапии аутоиммунных заболеваний, в частности, ревматоидного артрита . Дендритные клетки могут подавлять аутореактивные T-клетки и тем самым препятствовать развитию аутоиммунного процесса . Нарушения в работе дендритных клеток связаны с развитием других аутоиммунных заболеваний, таких как системная красная волчанка , болезнь Крона и язвенный колит , а также аллергии .
Дендритные клетки сами могут подвергаться злокачественному перерождению. В частности, при бластической неоплазии плазмоцитоидных дендритных клеток злокачественные плазмоцитоидные дендритные клетки проникают в кожу , костный мозг, нервную ткань и другие ткани и органы . Встречается также лейкоз , вызванный злокачественным перерождением плазмоцитоидных дендритных клеток. В этом случае злокачественные плазмоцитоидные дендритные клетки в большом количестве (минимум 2 % всех ядерных клеток крови ) присутствуют в кровотоке .
Дендритные клетки активизируются не только патогенами, их можно активировать искусственными стимуляторами. Пробиотический коктейль VSL#3 способен изменять экспрессию антигена и создание цитокинов. В высоких дозах пробиотик также повышает экспрессию молекул CD80, CD86, CD40 и MHC. С помощью пробиотиков можно регулировать количество дендритных клеток, таким образом способствуя лечению кишечных заболеваний .
В 2018 году удалось осуществить прямое перепрограммирование фибробластов в антигенпрезентирующие дендритные клетки, в ходе которого из фибробластов человека и мыши были получены дендритные клетки, представляющие антигены. В исследовании использовалась искусственная экспрессия факторов транскрипции для перепрограммирования дифференцированных клеток в сторону плюрипотентности . Путём комбинаций из 18 факторов транскрипции , экспрессируемые в клетках, были выявлены факторы транскрипции PU.1 , IRF8 и BATF3, нужные для перепрограммирования как мышиных, так и человеческих фибробластов в индуцированные дендритные клетки. Как и обычные дендритные клетки, они могут выделять цитокины, поглощать и презентировать антигены. Мышиные индуцированные дендритные клетки способны перекрёстно представлять антигены CD8+ Т-клеткам .
Дендритные клетки являются одной из мишеней вируса иммунодефицита человека . Одной из молекул, содействующих проникновению ВИЧ внутрь дендритных клеток, является поверхностный белок DC-SIGN . В дендритные клетки способны проникать и некоторые другие вирусы, например, SARS , также использующий DC-SIGN для проникновения в дендритные клетки .

Впервые дендритные клетки были обнаружены в 1868 году Паулем Лангергансом и впоследствии названы клетками Лангерганса в честь него . Лангерганс открыл дендритные клетки, будучи студентом университета, с помощью метода для выявления не пигментных клеток эпидермиса , который был разработан Юлиусом Конгеймом . Однако функции обнаруженных им отростчатых клеток Лангерганс не установил .
В 1957 году Фрэнк Макфарлейн Бёрнет предложил теорию клональной селекции , постулирующую, что при иммунном ответе активируются и делятся только те T-лимфоциты, T-клеточный рецептор которых связывает антиген . T-клетки неспособны реагировать на антигены самостоятельно, однако посредник, который представлял бы антигены T-клеткам, оставался неизвестным . В начале 1960-х годов было сформулировано представление о A-клетках — малочисленных клетках, перерабатывающих антиген и производящих его презентацию T-лимфоцитам. В 1966 году Роберт Мишелл и Ричард Даттон показали, что добиться активации T-клеток можно, добавив к ним суспензию из клеток селезёнки . В 1973 году Ральф Стайнман и применили термин «дендритные клетки» по отношению к отростчатым клеткам, располагающимся в лимфоидных органах. В 2011 году Стайнману была присуждена Нобелевской премии по физиологии или медицине за открытие дендритных клеток . К концу 1980-х годов были накоплены данные, свидетельствующие, что именно дендритные клетки являются «профессиональными» антигенпрезентирующими клетками .
| Органы | |||||
|---|---|---|---|---|---|
|
Врождённый
иммунитет |
|
||||
|
Приобретённый
иммунитет |
|
||||
| Медиаторы | |||||
| Иммунный ответ | |||||
|
Иммунологическая
толерантность |
|||||
| Нарушения | |||||
|
Лимфоцитарная
адаптивная иммунная система
и
комплемент
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Лимфоидные |
|
||||||||
| Лимфоциты | |||||||||
| Вещества | |||||||||